Есть ли у рыбы сердце? Рыбалка на гольяна – какими снастями следует заранее запастись. Виды красной рыбы
Не всякий ученый обладает цепким вниманием к предметам, казалось бы, давно известным и изученным. Зато именно таким людям дано совершать удивительные открытия.
К примеру, некто Томас Суини, обыкновенный учитель, преподававший в заштатном колледже где-то в американской глубинке, вовсе не являлся особым авторитетом в сфере биологии морских существ. Скорее, он был просто дилетантом, любителем, искренне увлеченным изучением жизни обитателей моря. Тем не менее, наблюдения, которые Т.Суини описал в своих письмах, адресованных некоторым специалистам в данной области, заставили этих авторитетных ученых глубоко задуматься и взглянуть на подводный мир другими глазами.
Такой вид симбиоза не был описан еще никем. Томасу Суини предстояло стать первооткрывателем.
Несмотря на излишне эмоциональное, непривычное для ученых, описание происходящего, отчет Томаса Суини был благосклонно принят его корреспондентами.
Вообще поведение рыб в таких подводных поликлиниках достойно удивления. Например, они строго соблюдают очередь, не допускают конфликтов, хотя рыбы здесь собираются самые разные.
Кроме того, пациенты полностью осознают пользу, приносимую им губанами, и даже помогают им себя очистить. Надо отметить, что некоторые рыбы приплывали на эти процедуры издалека, в обычной своей жизни редко посещая кораллы. Каким-то образом местонахождение такой поликлиники стало известно и им.
 Рыбы — строго организованные существа.
Рыбы — строго организованные существа. Но не стоит забывать и о том, что естественные законы бытия никто не отменял. В самих лечебных центрах соблюдается перемирие, но за границами рифа хищные рыбы вспоминают о своих пищевых предпочтениях, и тут прочим рыбкам приходится быть настороже.
Кстати говоря, мы часто находимся во власти стереотипов, даже если дело касается наших представлений о животных. К примеру, мурены в общепринятом представлении - злобные, наводящие ужас чудовища. Трудно себе представить, что кто-то может отзываться о них, как об очень деликатных существах, и даже называть их своими друзьями. А между тем, знаменитая женщина-фотограф В.Тэйлор, известная своими удивительными работами, посвященными жизни подводного мира, очень тепло вспоминает о своей дружбе с двумя муренами. Они ели у нее из рук, и она даже придумала им клички. Эти животные вовсе не были агрессивными просто так, а только в ответ на провокацию или в виде реакции на испуг. В общем-то, это не отличает их от прочих живых существ на земле или в воде.

Как оказалось, человек может наладить отношения и с муренами. В.Тэйлор, например, удавалось даже взять мурену в руки и поднять ее на поверхность воды. А играть и плавать с - это удовольствие вообще особого рода.
Конечно, далеко не каждый человек может подружиться с муреной или с гигантской мантой, заслужившей прозвище морского дьявола. Игры с этими чудовищами могут быть небезопасны, ведь вес манты может превышать тысячу килограмм. Человек по сравнению с этим гигантом — весьма маленькое и хрупкое создание. Тем не менее, истории о злонамеренности и кровожадности мант, которые утаскивают людей на дно, чтобы раздавить их, сильно преувеличены.
 Рыбы — существа, живущие по своим собственным законам.
Рыбы — существа, живущие по своим собственным законам. Валери Тэйлор говорит совершенно противоположные вещи. С ее точки зрения манты - вполне разумные и совсем не агрессивные создания, которые не собираются на вас нападать, если вы их не спровоцируете. При этом игра вовсе не считается провокацией! Играть манты любят и к прикосновениям относятся очень положительно. Кроме того, они обладают и своеобразной красотой и, несмотря на огромную массу тела, довольно изящно двигаются.
Ни наружного уха, ни барабанной перепонки у рыб нет. Звуковые волны передаются непосредственно через ткани. Лабиринт рыб служит одновременно и органом равновесия. Боковая линия дает возможность рыбе ориентироваться, чувствовать течение воды или приближение в темноте различных предметов. Органы боковой линии расположены в канале, погруженном в кожу, который сообщается с внешней средой при помощи отверстий в чешуе. В канале имеются нервные окончания.
Органы слуха рыб тоже воспринимают колебания водной среды, но только более высокочастотные, гармонические или звуковые. Устроены они у них более просто, чем у других животных.
Нет у рыб ни наружного, ни среднего уха: они обходятся без них в силу более высокой проницаемости воды для звука. Есть лишь перепончатый лабиринт, или внутреннее ухо, заключенное в костной стенке черепа.
Рыбы слышат, и притом отлично, так что рыболову во время уженья надо соблюдать полную тишину. Между прочим, это стало известно совсем недавно. Каких-нибудь 35-40 лет назад думали, что рыбы глухи.
По чувствительности на первый план зимой выступают слух и боковая линия. Здесь надо отметить, что внешние звуковые колебания и шумы сквозь ледяной и снежный покров в гораздо меньшей степени проникают в среду обитания рыб. В воде подо льдом устанавливается почти абсолютная тишина. И в таких условиях рыба в большей степени полагается на свой слух. Орган слуха и боковая линия помогают рыбе определять места скопления мотыля в донном грунте по колебаниям этих личинок. Если учесть также, что звуковые колебания затухают в воде в 3,5 тысячи раз медленнее, чем в воздухе, то становится ясно, что рыбы способны улавливать движения мотыля в донном грунте на значительном расстоянии.
Зарывшись в слой ила, личинки укрепляют стенки ходов затвердевающими выделениями слюнных желез и совершают в них волнообразные колебательные движения своим телом (рис.) , продувая и очищая свое жилище. От этого в окружающее пространство излучаются акустические волны, они-то и воспринимаются боковой линией и слухом рыб.
Таким образом, чем больше мотыля находится в донном грунте, тем больше исходит от него акустических волн и тем легче рыбе обнаружить самих личинок.
Уши у рыб есть! утверждает Юлия Сапожникова, научный сотрудник лаборатории ихтиологии. Только у них нет наружного уха, той самой ушной раковины, которую мы привыкли видеть у млекопитающих.
У некоторых рыбок нет уха, в котором были бы слуховые косточки молоточек, наковальня и стремечко также составляющие человеческого уха. Зато у всех рыб есть внутреннее ухо, и оно очень интересно устроено.
Рыбьи уши настолько малы, что умещаются на крошечных металлических "таблеточках", десяток которых свободно разместится на человеческой ладони.
На различные части внутреннего ушка рыбок наносится золотое напыление. Потом эти позолоченные рыбьи уши исследуют на электронном микроскопе. Только золотое напыление позволяет человеку увидеть детали внутреннего уха рыб. В золотой оправе их даже можно сфотографировать!
Камешек (отолит) под воздействием гидродинамических и звуковых волн совершает колебательные движения, а тончайшие сенсорные волоски улавливают их и передают сигналы головному мозгу.
Так рыбка различает звуки.
Ушной камешек оказался очень интересным органом. Например, если его расколоть, то можно на сколе увидеть кольца.
Это годовые кольца, точно такие есть на спиле деревьев. Поэтому по кольцам на ушном камешке, как по кольцам на чешуйках, можно определить, сколько рыбе лет.
Органы слуха и равновесия у рыб представлены внутренним ухом, наружное ухо у них отсутствует. Внутреннее ухо состоит из трех полукружных каналов с ампулами, овального мешочка и круглого мешочка с выступом (лагеной) . Рыбы единственные позвоночные с двумя или тремя парами отолитов, или ушных камешков, которые помогают поддерживать определенное положение в пространстве. У многих рыб имеется связь внутреннего уха с плавательным пузырем посредством цепи специальных косточек (веберов аппарат карповых, вьюновых и сомовых рыб) или с помощью доходящих до слуховой капсулы идущих вперед отростков плавательного пузыря (сельди, анчоусы, треска, многие морские караси, каменные окуни).
Способны ли рыбы слышать?
Поговорка «нем как рыба», с научной точки зрения давно утратило свою актуальность. Доказано, что рыбы умеют не только сами издавать звуки, но и слышать их. В течение долгого времени велись споры вокруг того, слышат ли рыбы. Сейчас ответ ученых известен и однозначен – рыбы не только обладают способностью слышать и имеют для этого соответствующие органы, но и сами посредством звуков в том числе могут между собой общаться.
Немного теории о сущности звука
Физиками давно установлено, что звук является ни чем иным, как цепочкой регулярно повторяющихся волн сжатия среды (воздушной, жидкой, твердой). Иначе говоря, звуки в воде являются столь же естественными, что и на ее поверхности. В воде звуковые волны, скорость которых обусловлена силой сжатия, могут распространяться различной частотой:
- большинство рыб воспринимает звуковые частоты в диапазоне 50-3000 Гц,
- вибрации и инфразвук, относящие к низкочастотным колебаниям до 16 Гц, воспринимают не все рыбы,
- способны ли рыбы воспринимать ультразвуковые волны, частота которых превышает 20000 Гц) – этот вопрос до конца еще не изучен, поэтому убедительные доказательства относительно наличия у подводных обитателей такой способности не получены.
Известно, что в воде звук распространяется вчетверо быстрее, нежели в воздухе или другой газообразной среде. Это – причина того, что звуки, которые поступают в воду извне, рыбы получают в искаженном виде. По сравнению с обитателями суши у рыб слух не столь острый. Однако эксперименты зоологов выявили очень интересные факты: в частности, некоторые виды раб умеют различать даже полутона.
Более подробно о боковой линии
Этот орган у рыб ученые относят к древнейшим сенсорным образованиям. Его можно считать универсальным, поскольку он выполняет не одну, а сразу несколько функций, обеспечивающих нормальную жизнедеятельность рыб.
Морфология латеральной системы не одинакова у всех видов рыб. Существуют ее варианты:
- Уже само расположение боковой линии на корпусе рыбы может относиться к специфичному признаку вида,
- Кроме того, известны виды рыб с двумя и более латеральными линиями по обеим сторонам,
- У костистых рыб боковая линия, как правило, проходит вдоль тела. У одних она непрерывная, у других – прерывистая и похожа на пунктир,
- У одних видов каналы латеральной линии спрятаны внутри кожи либо проходят открыто по поверхности.
Во всем остальном строение этого сенсорного органа у рыб идентично и функционирует он у всех видов рыб одинаково.
Этот орган реагирует не только на сжатие воды, но и на иные раздражители: электромагнитные, химические. Главную роль в этом играют невромасты, состоящие из, так называемых, волосковых клеток. Сама же структура невромастов это – капсула (слизистая часть), в которую и погружены собственно волоски чувствительных клеток. Поскольку сами невромасты закрыты, с внешней средой они соединены через микроотверстия в чешуе. Как мы знаем, невромасты бывают и открытым. Эти характерны для тех видов рыб, у которых каналов боковой линии заходят на голову.
В ходе многочисленных опытов, проводимых ихтиологами в разных странах было доподлинно установлено, что латеральная линия воспринимает низкочастотные колебания, причем, не только звуковые, но волны от движения других рыб.
Как органы слуха предупреждают рыб об опасности
В живой природе, как, в прочем, и в домашнем аквариуме, рыбы предпринимают адекватные меры, заслышав самые отдаленные звуки опасности. Пока шторм в этом районе моря или океана еще только зарождается, рыбы загодя меняют свое поведение – одни виды, опускаются на дно, где колебания волн наименьшие; другие мигрирую в спокойные локации.
Нехарактерные колебания воды расцениваются обитателями морей, как приближающаяся опасности и не отреагировать на нее они не могут, поскольку инстинкт самосохранения свойствен всему живому на нашей планете.
В реках поведенческие реакции рыб могут быть иными. В частности, при малейшем волнении воды (от лодки, например) рыба перестает есть. Это спасает ее от риска попасть на крючок к рыбаку.
Орган слуха рыб представлен только внутренним ухом и состоит из лабиринта, включающего преддверие и три полукружных канала, расположенных в трех перпендикулярных плоскостях. В жидкости, находящейся внутри перепончатого лабиринта, имеются слуховые камешки (отолиты) , колебания которых воспринимаются слуховым нервом. Ни наружного уха, ни барабанной перепонки у рыб нет. Звуковые волны передаются непосредственно через ткани. Лабиринт рыб служит одновременно и органом равновесия. Боковая линия дает возможность рыбе ориентироваться, чувствовать течение воды или приближение в темноте различных предметов. Органы боковой линии расположены в канале, погруженном в кожу, который сообщается с внешней средой при помощи отверстий в чешуе. В канале имеются нервные окончания. Органы слуха рыб тоже воспринимают колебания водной среды, но только более высокочастотные, гармонические или звуковые. Устроены они у них более просто, чем у других животных. Нет у рыб ни наружного, ни среднего уха: они обходятся без них в силу более высокой проницаемости воды для звука. Есть лишь перепончатый лабиринт, или внутреннее ухо, заключенное в костной стенке черепа. Рыбы слышат, и притом отлично, так что рыболову во время уженья надо соблюдать полную тишину. Между прочим, это стало известно совсем недавно. Каких-нибудь 35-40 лет назад думали, что рыбы глухи. По чувствительности на первый план зимой выступают слух и боковая линия. Здесь надо отметить, что внешние звуковые колебания и шумы сквозь ледяной и снежный покров в гораздо меньшей степени проникают в среду обитания рыб. В воде подо льдом устанавливается почти абсолютная тишина. И в таких условиях рыба в большей степени полагается на свой слух. Орган слуха и боковая линия помогают рыбе определять места скопления мотыля в донном грунте по колебаниям этих личинок.
Есть ли у рыб слух?
Если учесть также, что звуковые колебания затухают в воде в 3,5 тысячи раз медленнее, чем в воздухе, то становится ясно, что рыбы способны улавливать движения мотыля в донном грунте на значительном расстоянии. Зарывшись в слой ила, личинки укрепляют стенки ходов затвердевающими выделениями слюнных желез и совершают в них волнообразные колебательные движения своим телом (рис.) , продувая и очищая свое жилище. От этого в окружающее пространство излучаются акустические волны, они-то и воспринимаются боковой линией и слухом рыб. Таким образом, чем больше мотыля находится в донном грунте, тем больше исходит от него акустических волн и тем легче рыбе обнаружить самих личинок.
внутренее только
2 раздел


КАК РЫБЫ СЛЫШАТ |
Как известно, долгое время рыб считали глухими.
После того как у нас и за рубежом по методу условных рефлексов ученые провели эксперименты (в частности, среди подопытных были караси, окуни, лини, ерши и другие пресноводные рыбы), было убедительно доказано, что рыбы слышат, были также определены границы органа слуха, его физиологические функции и физические параметры.
Слух наряду со зрением - важнейший из чувств дистанционного (не контактного) действия, с его помощью рыбы ориентируются в окружающей среде. Без знания свойств слуха рыб нельзя до конца понять, каким образок поддерживается связь особей в косяке, как относятся рыбы к орудиям лова, каковы взаимоотношения хищника и жертвы. Прогрессирующей бионике необходим багаж накопленных фактов по строению и работе органа слуха у рыб.
Наблюдательные и смекалистые рыбаки-любители уже давно извлекали пользу из способности некоторых рыб слышать шум. Так родился способ ловли сомов на «клок». В насадке употребляют и лягушку; стремясь освободиться, лягушка, подгребая лапками, создает шум, хорошо знакомый сому, который часто оказывается тут как тут.
Итак, рыбы слышат. Давайте посмотрим на их орган слуха. У рыб не найти того, что называют наружным отделом органа слуха или ушами. Почему?
В начале этой книги мы упоминали о физических свойствах воды как прозрачной для звука акустической среды. Как бы пригодилась обитателям морей и озер способность навострять уши, подобно лосю или рыси, чтобы уловить далекий шорох и своевременно засечь крадущегося врага. Да вот незадача - оказывается, иметь уши не экономно для движения. Разглядывали щуку? Все ее точеное тело приспособлено для стремительного разгона и броска - ничего лишнего, что затрудняло бы движение.
Нет у рыб и так называемого среднего уха, свойственного наземным животным. У наземных животных аппарат среднего уха выполняет роль миниатюрного и просто устроенного приемо-передаточного преобразователя звуковых колебаний, осуществляющего свою paботу посредством барабанной перепонки и слуховых косточек. Эти «детали», слагающие конструкцию среднего уха наземных животных, у рыб имеют другое назначение, другое строение, другое название. И не случайно. Наружное и среднее ухо с его барабанной перепонкой биологически не оправдано в условиях больших, быстро нарастающих с глубиной давлений плотной массы воды. Интересно отметить, что у водных млекопитающих - китообразных, предки которых покинули сушу и вернулись в воду, барабанная полость не имеет выхода наружу, так как наружный слуховой проход либо заращен, либо перекрыт ушной пробкой.
И все-таки у рыб есть орган слуха. Вот его схема (см. рисунок). Природа позаботилась о том, чтобы этот весьма хрупкий, тонко устроенный орган был достаточно защищен - этим она как бы подчеркнула его значимость. (И у нас с вами внутреннее ухо защищает особо толстая кость). Вот лабиринт 2. С ним связана слуховая способность рыб (полукружные каналы - анализаторы равновесия). Обратите внимание на отделы, обозначенные цифрами 1 и 3. Это лагена (lagena) и саккулюс (sacculus) - слуховые приемники, рецепторы, воспринимающие звуковые волны. Когда в одном из опытов у гольянов с выработанным пищевым рефлексом на звук удалили нижнюю часть лабиринта - саккулюс и лагену, - они перестали отвечать на сигналы.
Раздражение по слуховым нервам передается в слуховой центр, расположенный в головном мозгу, где и происходят не постигнутые пока процессы превращения поступившего сигнала в образы и формирование ответной реакции.
Имеется два основных типа слуховых органов рыб: органы без связи с плавательным пузырем и органы, составной частью которых является плавательный пузырь.
Плавательный пузырь соединяется с внутренним ухом с помощью веберова аппарата - четырех пар подвижно сочлененных косточек. И хотя среднего уха у рыб нет, у некоторых из них (карповых, сомовых, харацинид, электрических угрей) есть его заменитель - плавательный пузырь плюс веберов аппарат.
До сих пор вы знали, что плавательный пузырь - это гидростатический аппарат, регулирующий удельный вес тела (а также то, что пузырь - необходимейший компонент полноценной карасевой ухи). Но об этом органе не лишне знать нечто большее. А именно: плавательный пузырь действует как приемник и преобразователь звуков (аналогично барабанной перепонке у нас). Вибрация его стенок передается через веберов аппарат и воспринимается ухом рыбы как колебания определенной частоты и интенсивности. С точки зрения акустики плавательный пузырь по существу представляет собой то же самое, что воздушная камера, помещенная в воду; отсюда - важные акустические свойства плавательного пузыря. Ввиду различия физических особенностей воды и воздуха акустический приемник
типа тонкой резиновой груши или плавательного пузыря, наполненный воздухом и помещенный в воду, при соединении с диафрагмой микрофона резко повышает его чувствительность. Внутреннее ухо рыбы и есть тот «микрофон», который работает в совокупности с плавательным пузырем. На деле это означает, что хотя раздел воды и воздуха в сильной степени отражает звуки, все же рыбы чувствительны к голосам и шуму с поверхности.
Всем известный лещ очень чуток в нерестовый период и боится малейшего шума. В старину во время нереста леща даже запрещалось звонить в колокола.
Плавательный пузырь не только повышает чувствительность слуха, но и расширяет воспринимаемый частотный диапазон звуков. В зависимости от того, сколько раз повторяются звуковые колебания за 1 секунду, измеряется частота звука: 1 колебание в секунду - 1 герц. Тикание карманных часов слышно в полосе частот от 1500 до 3000 герц. Для ясной, разборчивой речи по телефону достаточен диапазон частот от 500 до 2000 герц. Так что с гольяном мы смогли бы поговорить по телефону, ибо эта рыба реагирует на звуки в диапазоне частот от 40 до 6000 герц. Но если бы к телефону «подошли» гуппи, они бы услышали лишь те звуки, которые лежат в полосе до 1200 герц. Гуппи лишены плавательного пузыря, и их слуховой аппарат не воспринимает более высокие частоты.
В конце прошлого века экспериментаторы подчас не учитывали способностей различных видов рыб воспринимать звуки в ограниченном частотном диапазоне и делали ошибочные выводы об отсутствии слуха у рыб.
С первого взгляда может показаться, что возможности слухового органа рыбы никак нельзя сравнивать с чрезвычайно чувствительным ухом человека, способным обнаружить звуки ничтожно малой интенсивности и различать звуки, частоты которых лежат в диапазоне от 20 до 20000 герц. Тем не менее рыбы прекрасно ориентируются в родной стихии, и ограниченная порой частотная избирательность оказывается целесообразной, ибо позволяет из потока шума выделять только те звуки, которые оказываются полезными для особи.
Если звук характеризуется какой-либо одной частотой - мы имеем чистый тон. Чистый беспримесный тон получают с помощью камертона или звукового генератора. Большинство окружающих нас звуков содержит смесь частот, комбинацию тонов и оттенков тонов.
Надежным признаком развитого острого слуха служит способность различать тона. Человеческое ухо способно различать около полумиллиона простых тонов, различных по высоте и громкости. А как у рыб?
Гольяны способны различать звуки разной частоты. Дрессированные на определенный тон, они могут запоминать этот тон и реагировать на него, спустя один - девять месяцев после дрессировки. Некоторые особи могут запоминать до пяти тонов, например «до», «ре», «ми», «фа», «соль», и если «пищевой» тон при дрессировке был «ре», то гольян способен отличить его от соседнего более низкого тона «до» и более высокого тона «ми». Более того, гольяны в интервале частот 400-800 герц способны различать звуки, отличные по высоте на половину тона. Достаточно сказать, что фортепьянная клавиатура, удовлетворяющая самому тонкому человеческому слуху, содержит 12 полутонов октавы (отношение частот, равное двум, в музыке называется октавой). Что ж, возможно гольяны также «не лишены» некоторой музыкальности.
По сравнению со «слухачом» гольяном макропод не музыкален. Однако и макропод различает два тона, если они отстоят один от другого на 1 1/3 октавы. Можно упомянуть об угре, который замечателен не только тем, что идет нереститься за тридевять морей, но и тем, что способен различать звуки, отличные по частоте на октаву. Вышесказанное об остроте слуха рыб и их способности запоминать тона, заставляет по-новому перечитать строки известного австрийского аквалангиста Г. Хасса: «Не менее трехсот больших серебристых звездчатых ставрид подплыло сплошной массой и начало кружить вокруг громкоговорителя. Они держались от меня на расстоянии около трех метров и плыли словно в большом хороводе. Вполне вероятно, что звуки вальса - это были «Южные розы» Иоганна Штрауса - не имели ничего общего с этой сценой, и только любопытство, в лучшем случае звуки, привлекли животных. Но впечатление вальса рыб было столь полным, что я передал позже в нашем фильме так, как наблюдал сам».
Попытаемся теперь разобраться подробнее - что такое чувствительность слуха рыб?
Мы видим в отдалении двух беседующих людей, видим мимику каждого из них, жестикуляцию, но совершенно не слышим их голосов. Поток звуковой энергии, притекающий в ухо, настолько мал, что не вызывает слухового ощущения.
В данном случае чувствительность слуха можно оценивать наименьшей силой (громкостью) звука, которую ухо улавливает. Она отнюдь не одинакова по всему диапазону воспринимаемых данной особью частот.
Наивысшая чувствительность к звукам у человека наблюдается в полосе частот от 1000 до 4000 герц.
Ручьевой голавль в одном из экспериментов наименьший по силе звук воспринимал на частоте 280 герц. На частоте 2000 герц слуховая чувствительность его понижалась вдвое. Вообще рыбы лучше слышат низкие звуки.
Разумеется, слуховую чувствительность замеряют от какого-то начального уровня, принимаемого за порог чувствительности. Поскольку звуковая волна достаточной интенсивности производит вполне ощутимое давление, условились наименьшую пороговую силу (или громкость) звука определять в единицах давления, которое она оказывает. Такой единицей служит акустический бар. Нормальное человеческое ухо начинает улавливать звук, давление которого превышает 0,0002 бара. Чтобы понять, насколько это ничтожная величина, поясним, что звук карманных часов, прижатых к уху, оказывает на барабанную перепонку давление, превышающее пороговое в 1000 раз! В очень «тихой» комнате уровень звукового давления превышает пороговый в 10 раз. Это значит, что наше ухо фиксирует звуковой фон, который мы порой сознательно не в состоянии оценить. Для сравнения заметим, что барабанная перепонка испытывает боль, когда давление превышает 1000 бар. Такой силы звук мы чувствуем, стоя неподалеку от стартующего реактивного самолета.
Все эти цифры и примеры чувствительности человеческого слуха мы привели только для того, чтобы сопоставить их со слуховой чувствительностью рыб. Но не случайно говорят, что всякое сравнение хромает.
Есть ли у рыб уши?
Водная среда и особенности строения слухового органа рыб вносят заметные поправки в сравнительные измерения. Однако в условиях повышенного давления окружающей среды чувствительность человеческого слуха также заметно снижается. Как бы то ни было, но у карликового сомика чувствительность слуха ничуть не хуже человеческой. Это кажется поразительным, тем более что у рыб во внутреннем ухе нет кортиева органа - чувствительнейшего, тончайшего «прибора», который у человека и является собственно органом слуха.
Все это так: рыба слышит звук, рыба отличает один сигнал от другого по частоте и интенсивности. Но всегда следует помнить, что слуховые способности рыб не одинаковы не только между видами, но и среди особей одного вида. Если еще можно говорить о каком-то «усредненном» человеческом ухе, то по отношению к слуху рыб какой бы то ни было шаблон не применим, ибо особенности слуха рыб - результат жизни в конкретной обстановке. Может возникнуть вопрос: каким образом рыба отыскивает источник звука? Недостаточно слышать сигнал, надо сориентироваться на него. Жизненно важно для карася, до которого дошел грозный сигнал опасности - звук пищевого возбуждения щуки, локализовать этот звук.
Большинство изученных рыб способно локализовать звуки в пространстве на расстояниях от источников, приблизительно равных длине звуковой волны; на больших расстояниях рыбы обычно утрачивают способность определять направление к источнику звука и совершают рыскающие, поисковые движения, которые можно расшифровать как сигнал «внимание». Такая специфичность действия механизма локализации объясняется независимой работой двух приемников у рыб: уха и боковой линии. Ухо рыбы работает часто в комбинации с плавательным пузырем и воспринимает звуковые колебания в широком диапазоне частот. Боковая линия фиксирует давление и механические смещения частиц воды. Как ни малы сами по себе механические смещения частиц воды, вызванные давлением звука, они должны быть достаточными, чтобы их отметили живые «сейсмографы» - чувствительные клетки боковой линии. По-видимому, рыба получает информацию о расположении источника низкочастотного звука в пространстве сразу по двум показателям: величине смещения (боковая линия) и величине давления (ухо). Были проведены специальные опыты по выяснению способности речных окуней обнаруживать источники подводных звуков, излучаемых посредством магнитофона и гидроизолированных динамических наушников. В воду бассейна проигрывали записанные перед тем звуки питания - захват и перетирание пищи окунями. Такого рода опыты в аквариуме сильно усложняются тем, что многократное эхо от стенок бассейна как бы размазывает и заглушает основной звук. Похожий эффект наблюдается в обширном помещении с низким сводчатым потолком. Тем не менее окуни показали способность направленно, с расстояния до двух метров обнаруживать источник звука.
Метод пищевых условных рефлексов помог установить в условиях аквариума, что караси и карпы также способны определять направление к источнику звука. Некоторые морские рыбы (ставриды, рулены, барабули) в опытах в аквариуме и в море обнаруживали местоположение источника звука с расстояния 4-7 метров.
Но условия, в которых ставится опыт по выяснению той или иной акустической способности рыб, еще не дают представления о том, каким образом осуществляется звуковая сигнализация у рыб в естественной обстановке, где высок окружающий шумовой фон. Звуковой сигнал, несущий полезную информацию, только тогда имеет смысл, когда доходит до приемника в неискаженном виде, и это обстоятельство не требует особых пояснений.
У подопытных рыб, в том числе у плотвы и речного окуня, содержавшихся в аквариуме небольшими стайками, вырабатывали условный пищевой рефлекс. Как вы успели заметить, пищевой рефлекс фигурирует во многих опытах. Дело в том, что рефлекс на кормление быстро вырабатывается у рыб, и он наиболее устойчив. Аквариумисты это хорошо знают. Кто из них не проделывал простенький опыт: подкармливая рыб порцией мотыля, постукивать при этом по стеклу аквариума. После нескольких повторений, заслышав знакомый стук, рыбки дружно устремляются «к столу» - у них выработался рефлекс питания на условный сигнал.
В вышеуказанном опыте подавались два типа условных пищевых сигналов: однотонный звуковой сигнал частотой 500 герц, ритмически излучаемый через наушник посредством звукового генератора, и шумовой «букет», состоящий из предварительно записанных на магнитофон звуков, возникающих при питании особей. Для создания шумовой помехи в аквариум с высоты вливали струйку воды. В создаваемом ею фоновом шуме, как показали замеры, присутствовали все частоты звукового спектра. Нужно было выяснить, в состоянии ли рыбы выделить пищевой сигнал и среагировать на него в условиях маскировки.
Оказалось, что рыбы способны выделять полезные для них сигналы из шума. Причем однотонный звук, подаваемый ритмически, рыбы четко опознавали даже тогда, когда струйка падающей воды «забивала» его.
Звуки шумового характера (шорохи, чавканье, шелест, журчанье, шипенье и т. п.) рыбы выделяют (как и человек) лишь в случаях, когда они превышают уровень окружающих шумов.
Этот и другие аналогичные опыты доказывают способность слуха рыб выделить жизненно важные сигналы из набора бесполезных для особи данного вида звуков и шумов, в изобилии присутствующих в естественных условиях в любом водоеме, в котором есть жизнь.
На нескольких страницах мы рассмотрели возможности слуха рыб. Любители аквариума при наличии простых и доступных приборов, о которых мы поведем речь в соответствующей главе, могли бы самостоятельно поставить некоторые несложные опыты: например, определение способности рыб ориентироваться на источник звука, когда тот имеет для них биологическое значение, или способности рыб выделять такие звуки на фоне прочих «бесполезных» шумов, или обнаружение границы слуха у того или иного вида рыбы и т. д.
Многое еще не известно, многое нужно понять в устройстве и работе слухового аппарата рыб.
Хорошо изучены звуки, издаваемые тресковыми и сельдями, а слух их не исследован; у других рыб как раз наоборот. Полнее исследованы акустические возможности представителей семейства бычков. Так, один из них, черный бычок, воспринимает звуки, не превышающие частоту 800-900 герц. Все, что выходит за это частотный барьер, бычка «не касается». Его слуховые возможности позволяют воспринять хриплое, низкое ворчание, издаваемое соперником посредством плавательного пузыря; это ворчание в определенной ситуации можно расшифровать как сигнал угрозы. Но вот высокочастотные компоненты звуков, возникающие при питании бычков, ими не воспринимаются. И выходит, что какому-нибудь хитрому бычку, если он желает наедине полакомиться добычей, прямой расчет питаться на чуть более высоких тонах - соплеменники (они же конкуренты) его не услышат и не найдут. Это конечно шутка. Но в процессе эволюции вырабатывались самые неожиданные приспособления, порождавшиеся необходимостью жить в сообществе и зависеть хищнику от его жертвы, слабой особи от ее более сильного конкурента и т. д. И преимущества, даже небольшие, в способах получения информации (тоньше слух, обоняние, острее зрение и т. п.) оборачивались для вида благом.
В следующей главе мы покажем, что звуковые сигналы имеют в жизни рыбьего царства такое большое значение, о котором совсем недавно и не подозревали.
Вода - хранительница звуков …………………………………………………………………………….. 9
Как рыбы слышат
…………………………………………………………………………………………….. 17
Язык без слов - язык эмоций………………………………………………………………………………. 29
«Немые» среди рыб? ……………………………………………………………………………………………. 35
Рыбье «эсперанто» ………………………………………………………………………………………………. 37
Клев на уду! ………………………………………………………………………………………………………… 43
Не трепыхаться: акулы близко! ……………………………………………………………………………… 48
О «голосах» рыб и о том, что под этим понимается
и что из этого следует …………………………………………………………………………………………… 52
Сигналы рыб, связанные с размножением …………………………………………………………….. 55
«Голоса» рыб при обороне и нападении ……………………………………………………………….. 64
Незаслуженно забытое открытие барона
Мюнхгаузена ………………………………………………………………………………………………………… 74
«Табель о рангах» в стае рыб …………………………………………………………………………………. 77
Акустические вехи на путях миграций …………………………………………………………………… 80
Плавательный пузырь совершенствует
сейсмограф ……………………………………………………………………………………………………………. 84
Акустика или электричество? ………………………………………………………………………………… 88
О практической пользе изучения рыбьих «голосов»
и слуха………………………………………………………………………………………………………………….. 97
«Простите, нельзя ли с нами поделикатнее..?» …………………………………………………………97
Рыбаки надоумили ученых; ученые идут дальше ……………………………………………………. 104
Репортаж из недр косяка ……………………………………………………………………………………….. 115
Акустические мины и рыбы-подрывники ……………………………………………………………… 120
Биоакустика рыб в резерве у бионики ……………………………………………………………………. 124
Самодеятельному охотнику за подводными
звуками …………………………………………………………………………………………………………………. 129
Рекомендуемая литература …………………………………………………………………………………….. 143
Как слышат рыбы. Устройство уха
Мыне находим у рыбы ни ушных раковин, ни ушных отверстий. Но это ещё не значит, что у рыбы нет внутреннего уха, ведь и у нас наружное ухо само не ощущает звуков, а только помогает звуку достигнуть настоящего слухового органа - внутреннего уха, которое помещается в толще височной черепной кости.
Соответственные органы у рыбы помещаются также в черепе, по бокам головного мозга. Каждый из них имеет вид неправильного пузырька, наполненного жидкостью (рис. 19).
Звук может передаваться такому внутреннему уху через кости черепа, а возможность такой передачи звука мы можем обнаружить и на собственном опыте (плотно заткнув уши, приблизьте к самому лицу карманные или наручные часы - и вы не услышите их тиканья; приложите потом часы к зубам - тиканье часов будет слышно совершенно ясно).
Однако едва ли возможно сомневаться, что первоначальной и основной функцией слуховых пузырьков, когда они сформировались у древних предков всех позвоночных, было ощущение вертикального положения и что в первую очередь они являлись для водного животного статическими органами, или органами равновесия, вполне аналогичными статоцистам других свободноплавающих водных животных, начиная уже с медуз.
Таково же их важное жизненное значение и для рыбы, которая, согласно закону Архимеда, в водной среде практически оказывается «невесомой» и не может ощущать силы земного притяжения. Но зато каждое изменение в положении тела рыба ощущает слуховыми нервами, идущими к её внутреннему уху.
Её слуховой пузырёк наполнен жидкостью, в которой лежат крошечные, но весомые слуховые косточки: перекатываясь по дну слухового пузырька, они и дают рыбе возможность постоянно чувствовать вертикальное направление и сообразно этому двигаться.
Вопрос о том, слышат ли рыбы, долго дискутировался. В настоящее время установлено, что рыбы слышат и сами издают звуки. Звук представляет собой цепочку регулярно повторяющихся волн сжатия газообразной, жидкой или твердой среды, т. е. в водной среде звуковые сигналы так же естественны, как и на суше. Волны сжатия водной среды могут распространяться с различной частотой. Низкочастотные колебания (вибрация или инфразвук) до 16Гц воспринимаются не всеми рыбами. Однако у некоторых видов инфразвуковая рецепция доведена до совершенства (акулы). Спектр звуковых частот, воспринимаемый большинством рыб, лежит в диапазоне 50-3000 Гц. Способность к восприятию рыбами ультразвуковых волн (свыше 20 000 Гц) до настоящего времени убедительно не доказана.
Скорость распространения звука в воде в 4,5 раза больше, чем в воздушной среде. Поэтому звуковые сигналы с берега доходят до рыб в искаженном виде. Острота слуха у рыб не так развита, как у наземных животных. Тем не менее у некоторых видов рыб в экспериментах наблюдаются довольно приличные музыкальные способности. Например, гольян при 400-800 Гц различает 1/2 тона. Возможности других видов рыб скромнее. Так, гуппи и угорь дифференцируют два различающихся на 1/2-1/4 октавы. Есть и совершенно бездарные в музыкальном отношении виды (беспузырные и лабиринтовые рыбы).
|
|

Рис. 2.18. Связь плавательного пузыря с внутренним ухом у разных видов рыб: а- сельдь атлантическая; б -треска; в — карп; 1 — выросты плавательного пузыря; 2- внутреннее ухо; 3 — головной мозг: 4 и 5-косточки Веберова аппарата; общий эндолимфатический проток
Острота слуха определяется морфологией акустико-латеральной системы, к которой помимо боковой линии и ее производных относят внутреннее ухо плавательный пузырь и Веберов аппарат (рис. 2.18).
И в лабиринте, и в боковой линии чувствительными клетками выступают так называемые волосатые клетки. Смещение волоска чувствительной клетки как в лабиринте, так и в боковой линии приводит к одинаковому результату-генерации нервного импульса, поступающего в один и тот же акустико-латеральный центр продолговатого мозга. Однако эти органы рецептируют и другие сигналы (гравитационное поле, электромагнитные и гидродинамические поля, а также механические и химические раздражители).
Слуховой аппарат рыб представлен лабиринтом, плавательным пузырем (у пузырных рыб), Веберовым аппаратом и системой боковой линии. Лабиринт. Парное образование — лабиринт, или внутреннее ухо рыб (рис. 2.19), выполняет функцию органа равновесия и слуха. Слуховые рецепторы в большом количестве присутствуют в двух нижних камерах лабиринта — лагене и утрикулюсе. Волоски слуховых рецепторов очень чувствительны к перемещению эндолимфы в лабиринте. Изменение положения тела рыбы в любой плоскости приводит к перемещению эндолимфы, по крайней мере, в одном из полукружных каналов, что раздражает волоски.
В эндолимфе саккулы, утрикулюса и лагены находятся отолиты (камешки), которые повышают чувствительность внутреннего уха.
|
|

Рис. 2.19. Лабиринт рыбы: 1-круглый мешочек (лагена); 2-ампула (утрикулюс); 3-саккула; 4-каналы лабиринта; 5- расположение отолитов
Их общее количество по три с каждой стороны. Они различаются не только расположением, но и размерами. Самый крупный отолит (камешек) находится в круглом мешочке — лагене.
На отолитах рыб хорошо заметны годовые кольца, по которым v некоторых видов рыб определяют возраст. Они также обеспечивают оценку эффективности маневра рыбы. При продольном, вертикальном, боковом и вращательном движениях тела рыбы происходят некоторое смещение отолитов и раздражение ими чувствительных волосков, что, в свою очередь, создает соответствующий афферентный поток. На них же (отолиты) ложатся и рецепция гравитационного поля, оценка степени ускорения рыбы при бросках.
От лабиринта отходит эндолимфатический проток (см. рис. 2.18,6), который у костистых рыб закрыт, а у хрящевых открыт и сообщается с внешней средой. Веберов аппарат. Он представлен тремя парами подвижно соединенных косточек, которые называются стапесом (контактирует с лабиринтом), инкусом и малеусом (эта кость соединена с плавательным пузырем). Кости Веберова аппарата являются результатом эволюционной трансформации первых туловищных позвонков (рис. 2.20, 2.21).
При помощи Веберова аппарата лабиринт контактирует с плавательным пузырем у всех пузырных рыб. Другими словами, Веберов аппарат обеспечивает связь центральных структур сенсорной системы с воспринимающей звук периферией.

Рис.2.20. Строение Веберова аппарата:
1- перилимфатический проток; 2, 4, 6, 8- связки; 3 — стапес; 5- инкус; 7- малеус; 8 — плавательный пузырь (римскими цифрами обозначены позвонки)
|
|

Рис. 2.21. Общая схема строения органа слуха у рыбы:
1 — головной мозг; 2 — утрикулюс; 3 — саккула; 4- объединительный канал; 5 — лагена; 6- перилимфатический проток; 7-стапес; 8- инкус; 9-малеус; 10- плавательный пузырь
Плавательный пузырь. Он является хорошим резонирующим устройством, своеобразным усилителем средне- и низкочастотных колебаний среды. Звуковая волна извне приводит к колебаниям стенки плавательного пузыря, которые, в свою очередь, приводят к смещению цепочки косточек Веберова аппарата. Первая пара косточек Веберова аппарата давит на мембрану лабиринта, вызывая смещения эндолимфы и отолитов. Таким образом, если проводить аналогию с высшими наземными животными, Веберов аппарат у рыб выполняет функцию среднего уха.
Однако не у всех рыб есть плавательный пузырь и Веберов аппарат. В этом случае рыбы проявляют низкую чувствительность к звуку. У беспузырных рыб слуховую функцию плавательного пузыря частично компенсируют воздушные полости, связанные с лабиринтом, и высокая чувствительность органов боковой линии к звуковым стимулам (волнам сжатия воды).
Боковая линия. Она является очень древним сенсорным образованием, которое и у эволюционно молодых групп рыб выполняет одновременно несколько функций. Принимая во внимание исключительное значение этого органа для рыб, позволим себе более подробно остановиться на его морфофункциональной характеристике. Разные экологические типы рыб демонстрируют различные варианты латеральной системы. Расположение боковой линии на теле рыб часто является видоспецифичным признаком. Есть виды рыб, у которых более чем одна боковая линия. Например, терпуг имеет по четыре боковых линии с каждой стороны, отсюда
происходит его второе название — "восьмилинейный хир". У большинства костистых рыб боковая линия тянется вдоль туловища (не прерываясь или прерываясь в отдельных местах), достигает головы, образуя сложную систему каналов. Каналы боковой линии расположены или внутри кожи (рис. 2.22), или открыто на ее поверхности.
Примером открытого поверхностного расположения невромастов — структурных единиц латеральной линии — является боковая линия у гольяна. Несмотря на очевидное разнообразие морфологии латеральной системы, следует подчеркнуть, что наблюдаемые различия касаются только макростроения этого сенсорного образования. Собственно рецепторный аппарат органа (цепочка невромастов) на удивление одинаков у всех рыб как и морфологическом, так и функциональном отношении.
Система боковой линии реагирует на волны сжатия водной среды, обтекающие потоки, химические раздражители и электромагнитные поля при помощи невромастов — структур, объединяющих несколько волосковых клеток (рис. 2.23).
|
|

Рис. 2.22. Канал боковой линии рыбы
Невромаст состоит из слизисто-студенистой части — капулы, в которую погружены волоски чувствительных клеток. Закрытые невромасты сообщаются с внешней средой небольшими прободающими чешую отверстиями.
Открытые невромасты характерны для каналов латеральной системы, заходящих на голову рыбы (см. рис. 2.23, а).
Канальные невромасты тянутся от головы до хвоста по бокам тела, как правило, в один ряд (у рыб семейства Hexagramidae шесть рядов и бол ее). Термин "боковая линия" в обиходе относится именно к канальным невромастам. Однако у рыб описаны и невромасты, отделенные от канальной части и имеющие вид самостоятельных органов.
Канальные и свободные невромасты, расположенные в разных частях тела рыбы, и лабиринт не дублируют, а функционально дополняют друг друга. Считается, что саккулюс и лагена внутреннего уха обеспечивают звуковую чувствительность рыб с большого расстояния, а латеральная система позволяет локализовать источник звука (правда уже вблизи источника звука).

2.23. Строение невромастарыбы: а — открытый; б — канальный
Заметное влияние на активность рыб и характер их поведения оказывают волны, возникающие на поверхности воды. Причинами данного физического явления служат многие факторы: движение крупных объектов (крупная рыба, птицы, животные), ветер, приливы, землетрясения. Волнение служит важным каналом информирования водных животных о событиях как в самом водоеме, так и за его пределами. Причем волнение водоема воспринимается и пелагическими, и донными рыбами. Реакция на поверхностные волны со стороны рыбы бывает двух типов: рыба опускается на большую глубину или перемешается на другой участок водоема. Стимулом, действующим на тело рыбы в период волнения водоема, является движение воды относительно тела рыбы. Перемещение воды при ее волнении рецептируется акустико-латеральной системой, причем чувствительность боковой линии к волнам чрезвычайно высока. Так, для возникновения афферентации от боковой линии достаточно смешения купулы на 0,1 мкм. При этом рыба способна очень точно локализовать как источник волнообразования, так и направление распространения волны. Пространственная диаграмма чувствительности рыб видоспецифична (рис. 2.26).
В экспериментах использовали искусственный волнообразователь как очень сильный раздражитель. При изменении его местоположения рыбы безошибочно находили очаг возмущения. Реакция на источник волн состоит из двух фаз.
Первая фаза — фаза замирания — является результатом ориентировочной реакции (врожденного исследовательского рефлекса). Продолжительность этой фазы определяется многими факторами, наиболее существенными из которых являются высота волны и глубина погружения рыбы. Для карповых рыб (карп, карась, плотва) при высоте волны 2-12 мм и погружении рыб на 20-140 мм ориентировочный рефлекс занимал 200-250 мс.
Вторая фаза — фаза движения — условно-рефлекторная реакция вырабатывается у рыб довольно быстро. Для интактных рыб достаточно от двух до шести подкреплений для ее возникновений у ослепленных рыб после шести сочетаний волнообразования пищевого подкрепления вырабатывался устойчивый поисковый пищедобывающий рефлекс.
Большей чувствительностью к поверхностной волне отличаются Мелкие пелагические планктонофаги, меньшей — крупные донные рыбы. Так, ослепленные верховки при высоте волны всего 1- 3 мм уже после первого предъявления стимула демонстрировали ориентировочную реакцию. Для морских донных рыб характерна чувствительность к сильному волнению на поверхности моря. На глубине 500 м их латеральная линия возбуждается, когда высота волны достигает 3 м и длины 100 м. Как правило, волны на поверхности моря порождают качку Поэтому при волнении в возбуждение приходит не только боковая линия рыбы, но и ее лабиринт. Результаты экспериментов по, казали, что полукружные каналы лабиринта реагируют на вращательные движения, в которые водяные потоки вовлекают тело рыбы. Утрикулюс рецептирует линейное ускорение, возникающее в процессе качки. Во время шторма меняется поведение как одиночных, так и стайных рыб. При слабом шторме пелагические виды в прибрежной зоне опускаются в придонные слои. При сильном волнении рыбы мигрируют в открытое море и уходят на большую глубину, где влияние волнения менее заметно. Очевидно, что сильное волнение оценивается рыбами как неблагоприятный или даже опасный фактор. Он подавляет пищевое поведение и вынуждает рыб совершать миграции. Алогичные изменения в пищевом поведении наблюдаются и у видов рыб, обитающих во внутренних водоемах. Рыболовы знают, что при волнении моря клев рыбы прекращается.
Таким образом, водоем, в котором обитает рыба, является источником разнообразной информации, передаваемой по нескольким каналам. Такая информированность рыбы о колебаниях внешней среды позволяет ей своевременно и адекватно реагировать на них локомоторными реакциями и изменением вегетативных функций.
Сигналы рыб. Очевидно, что рыбы сами являются источником разнообразных сигналов. Они издают звуки в диапазоне частот от 20 Гц до 12 кГц, оставляют химический след (феромоны, кайромоны), имеют собственные электрические и гидродинамические поля. Акустические и гидродинамические поля рыбы создают различными способами.
Издаваемые рыбами звуки довольно разнообразны, однако из-за низкого давления зафиксировать их можно лишь при помощи специальной высокочувствительной техники. Механизм формирования звуковой волны у разных видов рыб может быть различным (табл. 2.5).
Звуки рыб видоспецифичны. Кроме того, характер звука зависят от возраста рыбы и ее физиологического состояния. Звуки, исходящие от стаи и от отдельных рыб, также хорошо различимы. Например, звуки, издаваемые лещом, напоминают хрипы. Звуковая картина стаи сельдей ассоциируется с писком. Морской петух Черного моря издает звуки, напоминающие кудахтанье курицы. Пресноводный барабанщик идентифицирует себя барабанной дробью. Плотва, вьюн, щитовка издают писки, доступные для восприятия невооруженным ухом.
Пока трудно однозначно охарактеризовать биологическое значение издаваемых рыбами звуков. Часть из них является шумовым фоном. Внутри популяций, стай, а также между половыми партнерами издаваемые рыбами звуки могут выполнять и коммуникативную функцию.
Шумопеленгация успешно применяется в промышленном рыболовстве.
есть ли у рыб уши?
Превышение звукового фона рыб над окружающими шумами составляет не более 15 дБ. Шумовой фон судна может десятикратно превышать рыбный звуковой пейзаж. Поэтому пеленг рыб возможен только с тех судов, которые могут работать в режиме "тишины", т. е. с заглушенными двигателями.
Таким образом, известное выражение "нем, как рыба" явно не соответствует действительности. Все рыбы имеют совершенный аппарат звуковой рецепции. Кроме того, рыбы являются источниками акустических и гидродинамических полей, которыми они активно пользуются для общения внутри стаи, обнаружения жертвы, предупреждения сородичей о возможной опасности и других целей.
Строение головного мозга костистых рыб
Головной мозг костистых рыб состоит из типичных для большинства позвоночных пяти отделов.
Ромбовидный мозг (rhombencephalon)
передним отделом заходит под мозжечок, а сзади без видимых границ переходит в спинной мозг. Чтобы рассмотреть передний отдел продолговатого мозга, необходимо отвернуть вперед тело мозжечка (у некоторых рыб мозжечок невелик и передний отдел продолговатого мозга хорошо виден). Крыша в этом отделе мозга представлена сосудистым сплетением. Под ним лежит большая расширенная на переднем конце и переходящая сзади в узкую медиальную щель, она является полостью Продолговатый мозг служит местом отхождения большинства головных нервов, а также проводящим путем, связывающим различные центры передних отделов головного мозга со спинным. Однако, слой белого вещества, покрывающий продолговатый мозг, у рыб довольно тонкий, так как туловище и хвост в значительной степени автономны - они осуществляют большую часть движений рефлекторно, не соотносясь с головным мозгом. В дне продолговатого мозга у рыб и хвостатых амфибий лежит пара гигантских маутнеровских клеток, связанных с акустико-латеральными центрами. Их толстые аксоны простираются вдоль всего спинного мозга. Локомоция у рыб осуществляется в основном за счёт ритмичных изгибов тела, которыми, по-видимому, управляют главным образом местные спинномозговые рефлексы. Однако общий контроль над этими движениями осуществляют маутнеровские клетки. В дне продолговатого мозга лежит дыхательный центр.
Рассматривая мозг снизу, можно различить места отхождения некоторых нервов. От латеральной стороны передней части продолговатого мозга отходят три круглых корешка. Первый, лежащий наиболее краниально, принадлежит V и VII нервам, средний корешок - только VII нерву, и, наконец, третий корешок, лежащий каудально, является VIII нервом. За ними, также от боковой поверхности продолговатого мозга, несколькими корешками отходят вместе IX и X пара. Остальные нервы тонки и обычно обрываются при препарировании.
Мозжечок (cerebellum) довольно хорошо развит, имеет округлую или вытянутую форму, он лежит над передней частью продолговатого мозга непосредственно за зрительными долями. Задним своим краем он прикрывает продолговатый мозг. Выступающая вверх часть является телом мозжечка (corpus cerebelli). Мозжечок является центром точной регуляции всех моторных иннервации, связанных с плаванием и схватыванием пищи.
Средний мозг (mesencephalon) - часть ствола головного мозга, пронизанная мозговым водопроводом. В его состав входят крупные, вытянутые продольно зрительные доли (они видны сверху).
Зрительные доли, или зрительная крыша (lobis opticus s. tectum opticus) - парные образования, отделенные друг от друга глубокой продольной бороздой. Зрительные доли являются первичными зрительными центрами, воспринимающими возбуждение. В них заканчиваются волокна зрительного нерва. У рыб этот отдел головного мозга имеет главенствующее значение, это центр, оказывающий основное влияние на деятельность организма. Серое вещество, покрывающее зрительные доли, имеет сложное слоистое строение, напоминающее строение коры мозжечка или полушарий
От вентральной поверхности зрительных долей отходят толстые зрительные нервы, перекрещивающиеся под поверхностью промежуточного мозга.
Если вскрыть зрительные доли среднего мозга, то можно увидеть, что в их полости от мозжечка отделяется складка, носящая название мозжечковой заслонки (valvule cerebellis). По бокам от нее в дне полости среднего мозга выделяются два бобовидных возвышения, называемых полулунными телами (tori semicircularis) и являющихся дополнительными центрами статоакустического органа.
Передний мозг (prosencephalon) менее развит, чем средний, он состоит из конечного и промежуточного мозга.
Части промежуточного мозга (diencephalon) лежат вокруг вертикально расположенной щели Боковые стенки желудочка - зрительные бугры или таламус (thalamus ) у рыб и амфибий имеют второстепенное значение (как координационные чувствительные и двигательные центры). Крыша третьего мозгового желудочка - надбугорье или эпиталамус - не содержит нейронов. В его составе находится переднее сосудистое сплетение (сосудистая покрышка третьего желудочка) и верхняя мозговая железа - эпифиз (epiphisis). Дно третьего мозгового желудочка - подбугорье или гипоталамус у рыб формирует парные вздутия - нижние доли (lobus inferior). Перед ними лежит нижняя мозговая железа - гипофиз (hypophisis). У многих рыб эта железа плотно входит в специальное углубление в дне черепа и обычно при препаровке обрывается; тогда хорошо видна воронка (infundibulum). перекрест зрительных нервов (chiasma nervorum opticorum).
у костистых рыб по сравнению с другими отделами мозга очень мал. Большинство рыб (кроме двоякодышащих и кистепёрых) отличает эвертированное (вывернутое) строение полушарий конечного мозга. Они как бы «вывернуты» вентро-латерально. Крыша переднего мозга не содержит нервных клеток, состоит из тонкой эпителиальной перепонки (pallium), которая при препаровке обычно снимается вместе с оболочкой мозга. В этом случае на препарате видно дно первого желудочка, разделенное глубокой продольной бороздой на два полосатых тела. Полосатые тела (corpora striatum1) состоят из двух отделов, что можно видеть при рассмотрении мозга сбоку. Фактически, эти массивные структуры содержат материал полосатых тел и коры довольно сложного строения.
Обонятельные луковицы (bulbus olfactorius) прилегают к переднему краю конечного мозга. От них вперед идут обонятельные нервы. У некоторых рыб (например, у трески) обонятельные луковицы вынесены далеко вперёд, в таком случае они соединяются с мозгом обонятельными трактами.
Черепномозговые нервы рыбы.
Всего от головного мозга рыбы отходит 10 пар нервов. В основном (и по наименованию и по функциям) они соответствуют нервам млекопитающих.
Строение головного мозга лягушки
Головной мозг лягушки, как и других амфибий, характеризуется следующими особенностями по сравнению с рыбами:
а) прогрессивным развитием головного мозга, выразившимся в обособлении парных полушарий продольной щелью и развитием в крыше мозга серого вещества древней коры (archipallium);
б) слабым развитием мозжечка;
в) слабой выраженностью изгибов головного мозга, благодаря чему промежуточный и средний отделы ясно видны сверху.

Ромбовидный мозг (rhombencephalon)
Продолговатый мозг (myelencephalon, medulla oblongata) , в который краниально переходит спинной мозг, отличается от последнего большей шириной и отхождением от его боковых поверхностей крупных корешков задних черепномозговых нервов. На дорсальной поверхности продолговатого мозга находится ромбовидная ямка (fossa rhomboidea), вмещающая четвёртый мозговой желудочек (ventriculus quartus). Сверху он прикрыт тонкой сосудистой покрышкой, которая удаляется вместе с мозговыми оболочками. По вентральной поверхности продолговатого мозга проходит вентральная щель - продолжение вентральной щели спинного мозга. В составе продолговатого мозга идут две пары тяжей (пучков волокон): нижняя пара, разделённая вентральной щелью - двигательные, верхняя пара - чувствительные. В продолговатом мозге расположены центры челюстного и подъязычного аппаратов, органа слуха, а также пищеварительной и дыхательной систем.
Мозжечок (cerebellum) располагается впереди ромбовидной ямки в виде высокого поперечного валика как выроста её передней стенки. Небольшие размеры мозжечка определяются небольшой и однообразной подвижностью амфибий - фактически, он состоит из двух небольших частей, тесно связанных с акустическими центрами продолговатого мозга (эти части сохраняются у млекопитающих как клочки мозжечка (flocculi)). Тело мозжечка - центр координации с другими отделами мозга - развито очень слабо.
Средний мозг (mesencephalon) при рассмотрении с дорсальной стороны представлен двумя типичными зрительными долями (lobus opticus s. tectum opticus) , имеющими вид парных яйцевидных возвышений, формирующих верхнюю и боковые части среднего мозга. Крыша зрительных долей образована серым веществом - несколькими слоями нервных клеток. Тектум у амфибий является наиболее значимым отделом головного мозга. В зрительных долях находятся полости, являющиеся боковыми ответвлениями мозгового (Сильвиева) водопровода (aquaeductus cerebri (Sylvii) , соединяющего четвёртый мозговой желудочек с третьим.
Дно среднего мозга формируют толстые пучки нервных волокон - ножки большого мозга (cruri cerebri), соединяющие передний мозг с продолговатым и спинным.
Передний мозг (prosencephalon) состоит из промежуточного и конечного мозга, лежащих последовательно.
сверху виден как ромб, острыми углами направленный в стороны.
Части промежуточного мозга лежат вокруг вертикально расположенной широкой щели третьего мозгового желудочка (ventriculus tertius). Боковые утолщения стенок желудочка - зрительные бугры или таламус (thalamus). У рыб и амфибий таламус имеет второстепенное значение (как координационные чувствительные и двигательные центры). Перепончатая крыша третьего мозгового желудочка - надбугорье или эпиталамус - не содержит нейронов. В его составе находится верхняя мозговая железа - эпифиз (epiphisis). У амфибий эпифиз уже выполняет роль железы, но не потерял ещё черты теменного органа зрения. Перед эпифизом промежуточный мозг покрыт перепончатой крышей, орально заворачивающейся внутрь и переходящей в переднее сосудистое сплетение (сосудистую покрышку третьего желудочка), а затем в концевую пластинку промежуточного мозга. Книзу желудочек сужается, формируя воронку гипофиза (infundibulum), каудовентрально к ней крепится нижняя мозговая железа - гипофиз (hypophisis). Впереди, на границе между дном конечного и промежуточного отделов головного мозга находится перекрест зрительных нервов (chiasma nervorum opticorum ). У амфибий большая часть волокон зрительных нервов не задерживается в промежуточном мозге, а идёт дальше - к крыше среднего мозга.
Конечный мозг (telencephalon) по своей длине почти равен длине всех остальных отделов головного мозга. Он состоит из двух частей: обонятельного мозга и двух полушарий, отделённых друг от друга сагиттальной (стреловидной) щелью (fissura sagittalis).
Полушария конечного мозга (haemispherium cerebri) занимают задние две трети конечного мозга и нависают над передней частью промежуточного мозга, частично прикрывая его. Внутри полушарий находятся полости - боковые мозговые желудочки (ventriculi lateralis), каудально сообщающиеся с третьим желудочком. В сером веществе полушарий головного мозга амфибий можно различить три участка: дорсомедиально находится старая кора или гиппокамп (archipallium, s. hippocampus), латерально - древняя кора (paleopallium) и вентролатерально - базальные ядра, соответствующие полосатым телам (corpora striata) млекопитающих. Полосатые тела и, в меньшей степени, гиппокамп являются коррелятивными центрами, последний связан с обонятельной функцией. Древняя кора является исключительно обонятельным анализатором. На вентральной поверхности полушарий заметны борозды, отделяющие полосатые тела от древней коры.
Обонятельный мозг (rhinencephalon) занимает переднюю часть конечного мозга и образует обонятельные доли (луковицы) (lobus olfactorius), спаянные посередине друг с другом. От полушарий они отделяются сбоку краевой ямкой. В обонятельные доли спереди входят обонятельные нервы.
От головного мозга лягушки отходят 10 пар черепномозговых нервов. Их образование, ветвление и зона иннервации принципиально не отличаются от таковых у млекопитающих
Головной мозг птиц.

Ромбовидный мозг (rhombencephalon) включает в себя продолговатый мозг и мозжечок.
Продолговатый мозг (myelencephalon, medulla oblongata) сзади непосредственно переходит в спинной мозг (medulla spinalis). Спереди он вклинивается между зрительными долями среднего мозга. Продолговатый мозг имеет толстое дно, в котором залегают ядра серого вещества - центры многих жизненно важных функций организма (в том числе равновесно-слуховой, соматические двигательные и вегетативные). Серое вещество у птиц покрыто толстым слоем белого, образованного нервными волокнами, соединяющими головной мозг со спинным. В дорсальной части продолговатого мозга имеется ромбовидная ямка (fossa rhomboidea), представляющая собой полость четвертого мозгового желудочка (ventriculus quartus). Крыша четвёртого мозгового желудочка образована перепончатой сосудистой покрышкой, у птиц она полностью прикрыта задним отделом мозжечка.
Мозжечок (cerebellum) у птиц крупный и представлен, практически, только червячком (vermis), расположенным над продолговатым мозгом. Кора (серое вещество, расположенное поверхностно) имеет глубокие борозды, значительно увеличивающие её площадь. Полушария мозжечка развиты слабо. У птиц хорошо развиты отделы мозжечка, связанные с мышечным чувством, в то время как отделы, отвечающие за функциональную связь мозжечка с корой полушарий, практически отсутствуют (они развиваются только у млекопитающих). На продольном разрезе хорошо видны полость желудочка мозжечка (ventriculus cerebelli), а также чередование белого и серого вещества, образующие характерный рисунок древа жизни (arbor vitae).
Средний мозг (mesencephalon) представлен двумя очень крупными, сдвинутыми вбок зрительными долями (lobus opticus s. tectum opticus). У всех позвоночных размер и развитие зрительных долей связан с размером глаз. Они хорошо видны сбоку и с вентральной стороны, тогда как с дорсальной стороны почти полностью прикрыты задними отделами полушарий. В зрительные доли у птиц приходят практически все волокна зрительного нерва, и зрительные доли остаются крайне важными частями головного мозга (однако у птиц со зрительными долями в значимости начинает соперничать кора полушарий). На сагиттальном разрезе видно, что по направлению вперёд полость четвёртого желудочка, суживаясь, переходит в полость среднего мозга - мозговой или силъвиев водопровод (aquaeductus cerebri). Орально водопровод переходит, расширяясь, в полость третьего мозгового желудочка промежуточного мозга. Условная передняя граница среднего мозга образована задней комиссурой (comissura posterior), хорошо заметной на сагиттальном разрезе в виде белого пятнышка.
В составе переднего мозга (prosencephalon) находятся промежуточный и конечный мозг.
Промежуточный мозг (diencephalon) у птиц снаружи виден только с вентральной стороны. Среднюю часть продольного разреза промежуточного мозга занимает узкая вертикальная щель третьего желудочка (ventriculus tertius). В верхней части полости желудочка видно отверстие (парное), ведущее в полость бокового желудочка - монроево (межжелудочковое) отверстие (foramen interventriculare).
Боковые стенки третьего мозгового желудочка образованы достаточно хорошо развитым у птиц таламусом (thalamus), степень развития таламуса связана со степенью развития полушарий. Он, не имея у птиц значения высшего зрительного центра, тем не менее, выполняет важные функции как двигательный коррелятивный центр.
В передней стенке третьего желудочка лежит передняя комиссура (comissura anterior), состоящая из соединяющих два полушария белых волокон
Дно промежуточного мозга именуют гипоталамусом (hypothalamus). При рассмотрении снизу видны боковые утолщения дна - зрительные тракты (tractus opticus). Между ними в передний конец промежуточного мозга входят зрительные нервы (nervus opticus), образующие зрительный перекрест (chiasma opticum). Задний нижний угол третьего мозгового желудочка соответствует полости воронки (infunbulum). Снизу воронка обычно прикрыта хорошо развитой у птиц подмозговой железой - гипофизом (hypophysis).
От крыши промежуточного мозга (эпиталамус (epithalamus) вверх отходит имеющая полость ножка пинеалъного органа. Выше находится и сам пинеалъный орган - эпифиз (epiphysis), он виден сверху, между задним краем больших полушарий и мозжечком. Передняя часть крыши промежуточного мозга образована сосудистым сплетением, заходящим в полость третьего желудочка.
Конечный мозг (telencephalon) у птиц состоит из больших полушарий (hemispherium cerebri), отделенных друг от друга глубокой продольной щелью (fissura interhemispherica). Полушария у птиц являются самыми крупными образованиями головного мозга, но структура их коренным образом отличается от таковой у млекопитающих. В отличие от мозга многих млекопитающих, сильно увеличенные полушария мозга птиц не несут борозд и извилин, поверхность их как с вентральной, так и с дорсальной стороны - гладкая. Кора в целом развита слабо, в первую очередь, в связи с редукцией органа обоняния. Тонкая медиальная стенка полушария переднего мозга в верхней части представлена нервным веществом старой коры (archipallium). Материал новой коры (слабо развита) (neopallium) вместе со значительной массой полосатых тел (corpus striatum) образует толстую боковую стенку полушария или боковой вырост, вдающийся в полость бокового желудочка. Поэтому полость бокового желудочка (ventriculus lateralis) полушария представляет собой узкую щель, расположенную дорсомедиально. У птиц в отличие от млекопитающих в полушариях значительного развития достигает не кора полушарий, а именно полосатые тела. Выявлено, что полосатые тела отвечают за врождённые стереотипные поведенческие реакции, в то время, как новая кора обеспечивает способности к индивидуальному обучению. У птиц некоторых видов обнаружено лучшее, чем в среднем, развитие участка новой коры - это, например, известные своими способностями к обучению вороны.
Обонятельные луковицы (bulbis olfactorius) располагаются на вентральной стороне переднего мозга. Они имеют небольшие размеры и примерно треугольную форму. Спереди в них входит обонятельный нерв.
Фото: Динары ВОРОНЦОВОЙ
Опытный аквариумист уверен: у некоторых – без сомнения
Магнитогорский биолог Владимир Пакулин занимается разведением рыб уже много лет.
Сколько он за свою жизнь сделал аквариумов, уже и не вспомнит. Устанавливал их у себя дома, помогал обустраивать рыбий быт у друзей, ставил и обслуживал аквариумы на предприятиях. Приходилось ухаживать за морскими обитателями, которые очень прихотливы, присматривал и за крокодилом, который оставил на память биологу шрам от укуса на руке.
Сегодня Владимир Леонидович на пенсии, подрабатывает сторожем в экологическом центре Дворца творчества детей и молодежи и продолжает заниматься любимым делом ребятишкам на радость, а себе в удовольствие.
Всех рыбок из дома он перевез в экоцентр: слишком много места занимают. Свой самый большой аквариум Владимир Пакулин планирует установить именно в экоцентре. Это будет гигантская чаша – две тонны воды, много растений, скала с водопадом, лягушки.
– Это будет даже не аквариум, а плюдариум, открытая емкость для содержания водных, погружных, прибрежных и наземных растений, животных, требующих высокой влажности в условиях, имитирующих тропический климат. В одной конструкции как бы совмещаются оранжерея, аквариум и террариум.
Много в экоцентре и обычных аквари-умов: с разноцветными рыбками, белыми лягушками, черепахами. Любимцы биолога – малавийские цихлиды. Это пресноводные рыбки, живущие в центральноафриканском озере Малави. Они живут только в этом озере, занимают все экологические ниши и инкубируют икру во рту. В природе цихлиды распространены очень широко. Они обитают в реках и озерах тропической части Центральной и Южной Америки, в водах Центральной Африки, Юго-Восточной Азии. Относятся рыбы этого семейства к отряду окунеобразных. Среди аквариумистов они пользуются популярностью за прекрасную окраску и оригинальную форму тела.
– Когда-то у меня был аквариум, где жило более 50 циклид. Они брали еду из рук, узнавали хозяина. Очень умные, я бы даже назвал их рыбами с интеллектом. Еще умным считаю астронотуса. Это крупная красивая рыбка со своеобразными повадками. Астронотусы тоже узнают хозяина, подплывают для общения, а постороннего человека пугаются – поворачиваются на бок и ложатся на дно.
Словно понимая, о чем мы говорим, несколько астронотусов, которые отлично себя чувствуют в экоцентре, «столпились»
за прозрачной стенкой аквариума и, казалось, внимательно слушали.
Есть в коллекции любителя аквариумов еще один занятный питомец – бойцовская рыбка петушок. Синий, с роскошным оперением, он плавал в небольшом круглом аквариуме.
– Эта рыбка очень неприхотлива, – рассказывал Владимир Леонидович. – Она дышит атмосферным воздухом. Для ее содержания не требуется компрессор, только смена воды. Но у нее особенность – нельзя сажать двух самцов вместе, они будут драться между собой. Еще одна странность этой рыбы – она может жить в круглом аквариуме.
А вот другие виды рыб этого не переносят. Нередко начинающие аквариумисты сажают рыбок в круглые аквариумы, и те по-рыбьи сходят с ума, начинают болеть, умирают. Всем рыбам нужны зрительная опора, ориентир на береговую линию. В аквариуме такой опорой служит задняя стенка. А вот петушок хорошо себя чувствует и без опоры.
Еще несколько советов дал опытный биолог Владимир Пакулин для начина-ющих аквариумистов:
– Чтобы в аквариуме не было войны между рыбками, надо знать, как рыба живет в природе, и создать ей необходимые условия: разместить водоросли, камешки, декор. Как это правильно сделать, можно прочитать в Интернете. Если условия содержания соблюдаются, каждый вид займет свою нишу. Аквариум – это многосемейный дом для рыб.
Если аквариумом занимается ребенок, идеальным вариантом будет золотая рыбка, яркая и неприхотливая. Но не забывайте устанавливать фильтр, компрессор и нагреватель. Питомцам можно дать клички, чтобы ребенок учился заботиться не об абстрактной рыбке, а о конкретном друге.
Начинающим не стоит заводить большой аквариум. Достаточно 40-литрового. Чем большее аквариум, тем труднее за ним ухаживать.
Чтобы рыбки не погибали, нужно правильно запустить их в новый аквариум. Для начала необходимо, чтобы готовый, со всем оборудованием, он постоял дней 10 без рыб. Затем нужно запустить самых дешевых рыбок, чтобы они создали комплекс микроорганизмов, который необходим более прихотливым рыбам. Через какое-то время, когда дешевые рыбки создадут благоприятный микроклимат, можно запускать уже дорогие виды рыб.





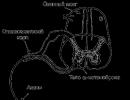
Есть ли у рыбы сердце?
Иногда нам очень трудно представить, что существа на нас совершенно не похожие могут иметь органы, очень напоминающие наши и функционирующие примерно так же. Многие думают, что раз рыба живет в воде и имеет холодную кровь, то у нее должны отсутствовать различные внутренние органы или какие-либо чувства. На самом же деле внутреннее строение рыбы очень похоже на строение высших, теплокровных животных.
Многие ученые считают, что это сходство доказывает то, что жизнь на суше появилась из моря! Рыбы дышат и переваривают пищу. У них есть нервная система, они чувствуют боль и физические неудобства. У них очень развито осязание. Они имеют вкусовые ощущения, а также очень чувствительную кожу. У них есть два маленьких органа обоняния в ноздрях, расположенных на голове. Даже уши у них есть, но они находятся внутри тела рыбы. Внешних органов слуха у рыбы нет. Глаза у рыб такие же, как и у позвоночных других видов, но имеют более простое строение.
Таким образом, вы можете видеть, что у рыбы имеются «системы», которые позволяют ей выполнять функции, сходные с функциями нашего организма. Давайте бегло рассмотрим лишь две из этих систем - пищеварения и кровообращения. Пища у рыбы проходит по пищеводу в брюшную полость, где находятся желудочные железы и где начинается переваривание пищи. Дальше она проходит в кишечник, где рассасывается, то есть поглощается кровью. Рыбы разных видов имеют и различные системы пищеварения, приспособленные к различным типам пищи - от растительной до другой рыбы. Но использует пищу рыба с такой же точно целью, что и мы: как источник энергии для жизни, роста и движения.
Система кровообращения рыбы разносит пищу и кислород во все внутренние органы. Насосом, регулирующим кровообращение рыбы, как и у человека, служит сердце. Сердце у рыбы находится за жабрами и чуть пониже их. Оно имеет три или четыре камеры, которые, как и у нас, ритмично сокращаются.
Существуют тысячи различных видов рыб, каждый из которых приспособлен к определенным жизненным условиям, но их внутренние органы, чувства и системы похожи на наши.