В каких органах находится мышечная ткань. Функции гладкой мышечной ткани
Подробности
Ганглии представляют собой скопления мультиполярных (один аксон и несколько дендритов) нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия.
Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны . В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля 1-го типа), чувствительные (клетки Догеля П-го типа) и ассоциативные (клетки Догеля Ш-го типа) . Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки 1-го типа называют длинноаксонными нейронами. Нейроны П-го типа - равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки П-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля Ш-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
Ацетилхолин
- никотиновые (блок кураре, гексаметоний), мускариновые (блок атропин) рецепторы. Активация рецепторов→генерирование ВПСП. Быстрый ВПСП (N-холиноцер)→открытие ионных каналов. Медленный ВПСП (М-холинорец)→подавление М-тока, обусловленного повышением К-проводимости.
Нейропептиды
– действуют как нейромодуляторы.
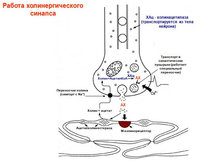
Энкефалины
, вещество Р, люлиберин, нейротензин, соматостатин – симп. ганглии (+Ach)
Катехоламины
(NA, дофамин) – нейромедиаторы мелких клеток с интенсивной флуоресценцией.
Нейропептид Y, соматостатин – симп. постганглионары.
Симпатические постганглионары: NA, АТФ, нейропептид У.
α1→инозотолтрифосфат, диацилглицерол. α2→активация G-белка, ↓цАМФ.
β→G-белок→AC→цАМФ

Исключения
: медиатор Ach, рецепторы мускариновые.
Парасимп. постганглионары: Ach, ВИП, NO, соматостатин, АТФ, опиоидные пептиды.
М1 (высокое сродство к пирензепину)-повышение секреции кислоты клетками желез желудка, М2 (низкое)- замедляют серд. ритм, секреция слезных и слюнные желез.
Разнообразное действие:
-Специфические втор. посредники: М2 может акт-ть IP3, а может инг-ть AC, уменьшая цАМФ.
-Действие на К и Са-каналы
-На эндотелии образуется NO→гуанилатциклаза→цГМФ→ цГМФ-зависимая протеинкиназа→расслабление глажких мышц.
Чувствительные нервные узлысодержат псевдоунипорлярные или биполярные афферентные нейроны и располагаются по ходу задних корешков спинного мозга (спинномозговые или спинальные ганглии) и черепномозговых нервов (V, VII, VIII, IX, X).
Спинномозговой узел (ganglion spinale) покрыт соединительноткан-ной капсулой. Внутри узла находятся группы псевдоуниполярных чувствительных нейронов, между которыми проходят пучки миелиновых волокон. Цитоплазма клеток содержит многочисленные митохондрии, цистерны гРЭПС, комплекс Гольджи, лизосомы. Тела нейронов окружены клетками - сателлитами (мантийные клетки) и соединительнотканной капсулой. Нейроны ганглия трех типов: малые, промежуточные и большие. Они различаются видами проводимых импульсов (тактильную чувствительность, пропреорецепцию, болевую, передают в ЦНС информацию о длине мышце и мышечном тонусе и т.д.). Они содержат нейромедиаторы: вещество Р, соматостатин и холецистокинин, глутамин, ВИП, гастрин. Их периферические отростки заканчиваются на периферии рецепторами. Центральные отростки (аксоны) входят в спинной мозг, образуя задние корешки спинного мозга и оканчиваются синапсами на вставочных нейронах и мотонейронах передних рогов спинного мозга.
Автономные (вегетативные) нервные узлы (ганглии) располагаются вдоль позвоночника в виде цепочки (паравентебральные ганглии) и впереди него (превентебральные ганглии), а также в стенке органов – сердце, пищеварительный тракт, мочевой пузырь и т.д. (интрамуральные ганглии) или вблизи поверхности органов (экстрамуральные ганглии).
К вегетативным ганглиям подходят преганглионарные волокна (миелиновы, содержащие отростки нервных клеток, тела которых лежат в ЦНС. Волокна сильно ветвятся и образуют синаптические окончания на клетках вегетативных ганглий.
Вегетативные ганглии по функциональному признаку и локализации разделяются на симпатические и парасимпатические.
Симпатические нервные узлы (паравертебральные и превертебральные) получают преганглионарные волокна от клеток, расположенных в вегетативных ядрах грудных и поясничных сегментов спинного мозга. Нейромедиатором преганглионарных волокон является ацетилхолин , а постганглионарных – норадреналин (исключение потовые железы и некоторые кровеносные сосуды, которые имеют хилинергическую симпатическую иннрвацию).В узлах выявляются также энкефалины, ВИП, вещество Р, соматостатин, холецистокенин.
Парасимпатические нервные узлы (интрамуральные, экстрамуральные или узлы головы) получают преганглионарные волокна от клеток, тела которых расположены в вегетативных ядрах продолговатого мозга, среднего мозга, и в кресцовом отделе спинного мозга. Эти волокна покидаю ЦНС в составе III, VII, IX и X пар черепномозговых нервов и передних корешков кресцовых сегментов спинного мозга. Нейромедиатором пре- и постганглионарных волокон является ацетилхолин, а также серотонин АТФ и др.
Болшинство органов получают как симпатическую, так и парасимпатическую иннервацию.
Строение симпатических и парасимпатических ганглиев в общем плане сходно. Вегетативные ганглии с поверхности покрыты соединительнотканной капсулой, которая, проникая внутрь узла, образует строму. Узлы состоят из мультиполярных нервных клеток, различных по форме и размерам и их отростков. Тела нейронов неправильной формы, с эсцентрично расположенными ядрами, окружены оболочками из глиальных клеток – сателлитов (мантийные глиоциты). Отростки клеток также покрыты глиальными клетками. Глиальная оболочка покрыта базальной мебраной, поверх которой находится соединительнотканная оболочка.
В симпатических ганглиях наряду с крупными клетками содержатся небольшие группы мелких клеток с интенсивно флюоресцирующими гранулами МИФ-клетки, и мелкие гранулосодержащие клетки (МСГ-клетки). В гранулах содержатся дофамин, серотонин или норадреналин. На МИФ клетках оканчиваются терминали преганглионарных волокон, при стимуляции которых выделяются медиаторы в периваскулярные пространства и в области синапсов на дендритах крупных клеток. МИФ-клетки обладают ингибирующим действием на эффекторные клетки.
Интрамуральные узлы - это нервные ганглии, расположенные внутри иннервируемых органов. Интрамуральные ганглии и связанные и ними проводящие пути обладают высокой автономией, сложностью организации и особенностью медиаторов, и в связи с этим многими авторами выделяют в самостоятельный метасимпатический отдел вегетативной нервной системы.
Наиболее наглядно строение интрамуральных ганглиев можно проследить на примере вегетативной иннервации пищеварительного тракта. В пищеварительной трубке содержится два крупных нервных сплетения: подслизистое – Мейснера , межмышечное – Ауэрбаха. Общее число нейронов в интрамуральных ганглиях выше, чем в спинном мозге, а по сложности их взаимодействия их сравнивают с микрокомпьютером.
В интрамуральных узлах описаны нейроны 3-х типов. Первые данные по гетерогенности нейронов пищеварительного тракта были получены Догелем. Основываясь на форме клеток и характере ветвления их отростков Догель выделил три типа нейронов.
1. длинноаксонные эфферентные нейроны (клетки Догеля I типа) численно преобладают. Это крупные или средние клетки, перикарионы уплощенной форммы с короткими дендритами и длинным аксоном, который направляется за пределы узла и оканчивается на клетках рабочего органа двигательным или секреторным окончанием.
Представляет собой систему тканей и органов, построенных из нервной ткани. В ней выделяют:
Центральный отдел: головной и спинной мозг
Периферический отдел: автономные и чувствительные ганглии, периферические нервы, нервные окончания.
Существует также подразделение на:
Соматический (анимальный, цереброспинальный) отдел;
Вегетативный (автономный) отдел: симпатическая и парасимпатическая части.
Нервную систему формируют следующие эмбриональные источники: нервная трубка, нервный гребень (ганглиозная пластинка) и эмбриональные плакоды. Тканевые элементы оболочек являются мезенхимными производными. На стадии замыкания нейропоров передний конец трубки значительно расширяется, боковые стенки утолщаются, образуя зачатки трех мозговых пузырей. Лежащий краниально пузырь образует передний мозг, средний пузырь - средний мозг, а из третьего пузыря, который переходит в закладку спинного мозга, развивается задний (ромбовидный) мозг. Вскоре после этого нервная трубка изгибается почти под прямым углом, и посредством борозд-сужений первый пузырь разделяется на конечный и промежуточный отделы, а третий мозговой пузырь - на продолговатый и задний отделы мозга. Производные среднего и заднего мозговых пузырей образуют ствол мозга и являются древними образованиями; в них сохраняется сегментарный принцип строения, который исчезает в производных промежуточного и конечного мозга. В последних концентрируются интегративные функции. Так формируются пять отделов мозга: конечный и промежуточный мозг, средний, продолговатый и задний мозг (у человека это происходит примерно в конце 4-й нед эмбрионального развития). Конечный мозг формирует два полушария большого мозга.
В эмбриональном гисто- и органогенезе нервной системы развитие разных отделов мозга происходит с разной скоростью (гетерохронно). Раньше формируются каудальные отделы центральной нервной системы (спинной мозг, ствол мозга); время окончательного формирования структур головного мозга сильно варьирует. В ряде отделов головного мозга это происходит после рождения (мозжечок, гиппокамп, обонятельная луковица); в каждом отделе мозга существуют пространственно-временные градиенты формирования нейронных популяций, которые образуют уникальную структуру нервного центра.
Спинной мозг представляет собой часть центральной нервной системы, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегмен-тарность. В боковых отделах нервной трубки быстро возрастает масса клеток, тогда как дорсальная и вентральная ее части не увеличиваются в объеме и сохраняют эпендимный характер. Утолщенные боковые стенки нервной трубки делятся продольной бороздой на дорсальную - крыльную, и вентральную - основную пластинку. На этой стадии развития в боковых стенках нервной трубки можно выделить три зоны: эпендиму, выстилающую центральный канал, промежуточную (плащевой слой) и маргинальную (краевую вуаль). Из плащевого слоя в дальнейшем развивается серое вещество спинного мозга, а из краевой вуали - его белое вещество. Нейробласты передних столбов дифференцируются в мотонейроны (двигательные нейроны) ядер передних рогов. Их аксоны выходят из спинного мозга и образуют передние корешки спинномозговых нервов. В задних столбах и промежуточной зоне развиваются различные ядра вставочных (ассоциативных) клеток. Их аксоны, поступая в белое вещество спинного мозга, входят в состав различных проводящих пучков. В задние рога входят центральные отростки чувствительных нейронов спинномозговых узлов.
Одновременно с развитием спинного мозга закладываются спинномозговые и периферические узлы автономной нервной системы. Исходным материалом для них служат стволовые клеточные элементы нервного гребня, которые путем дивергентной дифференцировки развиваются в нейробла-стическом и глиобластическом направлениях. Часть клеток нервного гребня мигрирует на периферию в места локализации узлов автономной нервной системы, параганглиев, нейроэндокринных клеток APUD-серии и хромаффинной ткани.
Периферическая нервная система.
Периферическая нервная система объединяет периферические нервные узлы, стволы и окончания.
Нервные ганглии (узлы) – структуры, образованные скоплениями нейронов вне ЦНС, - разделяются на чувствительные и автономные (вегетативные). Чувствительные ганглии содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются преимущественно по ходу задних корешков спинного мозга (чувствительные узлы спинномозговых нервов) и некоторых черепно-мозговых нервов. Чувствительные ганглии спинномозговых нервов имеют веретеновидную форму и покрыты капсулой из плотной волокнистой соединительной ткани. По периферии ганглия находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды. Автономные нервные ганглии образованы скоплениями мультиполярных нейронов, на которых многочисленные синапсы образуют преганглионарные волокна – отростки нейронов, чьи тела лежат в ЦНС.
Нерв. Строение и регенерация. Спинномозговые ганглии. Морфофункциональная характеристика.
Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками миелиновых и безмиелиновых волокон, которые объединены соединительнотканными компонентамии (оболочками): эндоневрием, периневрием и эпиневрием. Большинство нервов являются смешанными, т.е. включают афферентные и эфферентные волокна.
Эндоневрий – тонкие прослойки рыхлой волокнистой соединительной ткани с мелкими кровеносными сосудами, окружающие отдельные нервные волокна и связывающие их в единый пучок. Периневрий – оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Он имеет пластинчатое строение и образов концентрическими пластами уплощённых фиброблатстоподобных клеток, связанных плотными и щелевыми соединениями. Между слоями клеток в пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна. Эпиневрий – наружная оболочка нерва, связывающая воедино пучки нервных волокон. Он состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды.
Спинной мозг. Морфофункциональная характеристика. Развитие. Строение серого и белого вещества. Нейронный состав.
Спинной мозг состоит из двух симметричных половин, отграниченных друг от друга спереди глубокой серединной щелью, а сзади – соединительнотканной перегородкой. Внутренняя часть органа темнее - это его серое вещество. На периферии спинного мозга располагается более светлое белое вещество. Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон и нейроглии. Основной составной частью серого вещества, отличающей его от белого, являются мультиполярные нейроны. Выступы серого вещества принято называть рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога. В процессе развития спинного мозга из нервной трубки образуются нейроны, группирующиеся в 10 слоях, или в пластинах. Для человека характерна
следующая архитектоникауказанных пластин: I-V пластины соответствуют задним рогам, VI-VII пластины - промежуточной зоне, VIII-IX пластины - передним рогам, X пластина - зона околоцентрального канала. Серое вещество мозга состоит из мультиполярных нейронов трех типов. Первый тип нейронов является филогенетически более древним и характеризуется немногочисленными длинными, прямыми и слабо ветвящимися дендритами (изодендритический тип). Второй тип нейронов имеет большое число сильно ветвящихся дендритов, которые переплетаются, образуя «клубки» (идиодендритический тип). Третий тип нейронов по степени развития дендритов занимает промежуточное положение между первым и вторым типами. Белое вещество спинного мозга представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга
Головной мозг. Источники развития. Общая морфофункциональная характеристика больших полушарий. Нейронная организация больших полушарий. Цито- и миелоархитектоника коры больших полушарий головного мозга. Возрастные изменения коры.
В головном мозге различают серое и белое вещество, но распределение этих двух составных частей здесь значительно сложнее, чем в спинном мозге. Большая часть серого вещества головного мозга располагается на поверхности большого мозга и в мозжечке, образуя их кору. Меньшая часть образует многочисленные ядра ствола мозга.
Строение . Кора большого мозга представлена слоем серого вещества. Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга.. Различные участки ее, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные
границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями. Развитие коры больших полушарий (неокортекса) человека в эмбриогенезе происходит из вентрикулярной герминативной зоны конечного мозга, где расположены малоспециализированные пролиферирующие клетки. Из этих клеток дифференцируются нейроциты неокортекса. При этом клетки утрачивают способность к делению и мигрируют в формирующуюся корковую пластинку. Вначале в корковую пластинку поступают нейроциты будущих I и VI слоев, т.е. наиболее поверхностного и глубокого слоев коры. Затем в нее встраиваются в направлении изнутри и кнаружи последовательно нейроны V, IV, III и II слоев. Этот процесс осуществляется за счет образования клеток в небольших участках вентрикулярной зоны в различные периоды эмбриогенеза (гетерохрон-но). В каждом из этих участков образуются группы нейронов, последовательно выстраивающихся вдоль одного или нескольких волокон
радиальной глии в виде колонки.
Цитоархитектоника коры большого мозга. Мультиполярные нейроны коры весьма разнообразны по форме. Среди них можно выделить пирамидные, звездчатые, веретенообразные, паукообразные и горизонтальные нейроны. Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоев: I - молекулярный, II - наружный зернистый, III - nuрамидных нейронов, IV - внутренний зернистый, V - ганглионарный, VI - слой полиморфных клеток. Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их нейриты проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Наружный зернистый слой образован мелкими нейронами, имеющими округлую, угловатую и пирамидальную форму, и звездчатыми нейроцитами. Дендриты этих клеток поднимаются в молекулярный слой. Нейриты или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя. Самый широкий слой коры большого мозга - пирамидный. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Нейрит пирамидной клетки всегда отходит от ее основания. Внутренний зернистый слой образован мелкими звездчатыми нейронами. В его состав входит большое количество горизонтальных волокон. Ганглионарный слой коры образован крупными пирамидами, причем область прецентральной извилины содержит гигантские пирамиды.
Слой полиморфных клеток образован нейронами различной формы.
Миелоархитектоника коры . Среди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной
нервной системы.
Возрастные изменения . На 1-м году жизни наблюдаются типизация формы пирамидных и звездчатых нейронов, их увеличение, развитие дендритных и аксонных арборизаций, внутриансамблевых связей по вертикали. К 3 годам в ансамблях выявляются «гнездные» группировки нейронов, более четко сформированные вертикальные дендритные пучки и пучки радиарных волокон. К 5-6 годам нарастает полиморфизм нейронов; усложняется система внутриансамблевых связей по горизонтали за счет роста в длину и разветвлений боковых и базальных дендритов пирамидных нейронов и развития боковых терминалей их апикальных дендритов. К 9-10 годам увеличиваются клеточные группировки, значительно усложняется структура короткоаксонных нейронов, и расширяется сеть аксонных коллатералей всех форм интернейронов. К 12-14 годам в ансамблях четко обозначаются специализированные формы пирамидных нейронов, все типы интернейронов достигают высокого уровня дифференцировки. К 18 годам ансамблевая организация коры по основным параметрам своей архитектоники достигает уровня таковой у взрослых.
Мозжечок. Строение и морфофункциональная характеристика. Нейронный состав коры мозжечка, глиоциты. Межнейронные связи.
Мозжечок . Представляет собой центральный орган равновесия и координации движений. Он связан со стволом мозга афферентными и эфферентными проводящими пучками, образующими в совокупности три пары ножек мозжечка. На поверхности мозжечка много извилин и бороздок, которые значительно увеличивают ее площадь. Борозды и извилины создают на разрезе
характерную для мозжечка картину «древа жизни». Основная масса серого вещества в мозжечке располагается на поверхности и образует его кору. Меньшая часть серого вещества лежит глубоко в белом веществе в виде центральных ядер. В центре каждой извилины имеется тонкая прослойка
белого вещества, покрытая слоем серого вещества - корой. В коре мозжечка различают три слоя: наружный - молекулярный, средний - ганглионарный слой, или слой грушевидных нейронов, и внутренний -зернистый. Ганглиозный слой содержит грушевидные нейроны. Они имеют нейриты, которые, покидая кору мозжечка, образуют начальное звено его эфферентных
тормозных путей. От грушевидного тела в молекулярный слой отходят 2-3 дендрита, которые пронизывают всю толщу молекулярного слоя. От основания тел этих клеток отходят нейриты, проходящие через зернистый слой коры мозжечка в белое вещество и заканчивающиеся на клетках ядер мозжечка. Молекулярный слой содержит два основных вида нейронов: кор-зинчатые и звездчатые. Корзинчатые нейроны находятся в нижней трети молекулярного слоя. Их тонкие длинные дендриты ветвятся преимущественно в плоскости, расположенной поперечно к извилине. Длинные нейриты клеток всегда идут поперек извилины и параллельно поверхности над грушевидными нейронами. Звездчатые нейроны лежат выше корзинчатых и эывают двух типов. Мелкие звездчатые нейроны снабжены тонкими короткими дендритами и слаборазветвленными нейритами, образующими синапсы. Крупные звездчатые нейроны имеют длинные и сильно разветвленные дендриты и нейриты. Зернистый слой. Первым типом клеток этого слоя можно считать зерновидные нейроны, или клетки-зерна. Клетка имеет 3-4 коротких дендрита,
заканчивающихся в этом же слое концевыми ветвлениями в виде лапки птицы. Нейриты клеток-зерен проходят в молекулярный слой и в нем делятся на две ветви, ориентированные параллельно поверхности коры вдоль извилин мозжечка. Вторым типом клеток зернистого слоя мозжечка являются тормозные большие звездчатые нейроны. Различают два вида таких клеток: с короткими и длинными нейритами. Нейроны с короткими нейритами лежат вблизи ганглионарного слоя. Их разветвленные дендриты распространяются в молекулярном слое и образуют синапсы с параллельными волокнами - аксонами клеток-зерен. Нейриты направляются в зернистый слой к клубочкам мозжечка и заканчиваются синапсами на концевых ветвлениях дендритов клеток-зерен.
Немногочисленные звездчатые нейроны с длинными нейритами имеют обильно ветвящиеся в зернистом слое дендриты и нейриты, выходящие в белое вещество. Третий тип клеток составляют веретеновидные горизонтальные клетки. Они имеют небольшое вытянутое тело, от которого в обе стороны отходят длинные горизонтальные дендриты, заканчивающиеся в ганглионарном и зернистом слоях. Нейриты же этих клеток дают коллатерали в зернистый слой и уходят в
белое вещество. Глиоциты . Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны. Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В ганглионарном слое между грушевидными нейронами лежат глиальные клетки с темными ядрами. Отростки этих клеток направляются к поверхности коры и образуют глиальные волокна молекулярного слоя мозжечка. Межнейрональные связи . Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами - моховидными и так называемыми лазящими волокнами. Моховидные волокна идут в составе оливомозжечкового и мостомозжечкового путей и опосредованно через клетки-зерна оказывают на грушевидные клетки возбуждающее действие.
Лазящие волокна поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям. Они пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами. Лазящие волокна передают возбуждение непосредственно грушевидным нейронам.
Автономная (вегетативная) нервная система. Общая морфофункциональная характеристика. Отделы. Строение экстрамуральных и интрамуральных ганглиев.
ВНС делится на симпатическую и парасимпатическую. Обе системы одновременно принимают участие в иннервации органов и оказывают на них противоположное влияние. Состоит из центральных отделов, представленных ядрами серого вещества головного и спинного мозга, и периферических: нервных стволов, узлов (ганглиев) и сплетений.
Интрамуральные ганглии и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена выделяют в самостоятельный метасимпатический отдел автономной НС. Выделяют нейроны трёх типов:
Длинноаксонные эфферентные нейроны (клетки I типа Догеля) с короткими дендритами и длинным аксоном, идущим за пределы узла к клеткам рабочего органа, на которых он образует двигательные или секреторные окончания.
Равноотросчатые афферентные нейроны (клетки II типа Догеля) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС.
Ассоциативные клетки (клетки III типа Догеля) – местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
В вегетативной нервной системе
различают центральные и периферические отделы. Центральные отделы симпатической нервной системы представлены ядрами боковых рогов тораколюмбального отдела спинного мозга. В парасимпатической нервной системе центральные отделы включают ядра среднего и продолговатого мозга, а также ядра боковых рогов сакрального отдела спинного мозга. Парасимпатические волокна краниобульбарного отдела выходят в составе III-й, VII-й, IX-й и Х-й пар черепных нервов.
Периферические отделы вегетативной нервной системы
образованы нервными стволами, ганглиями и сплетениями.
Вегетативные рефлекторные дуги начинаются чувствительным нейроном, тело которого лежит в спинномозговом узле (ганглии), как и в соматической рефлекторной дуг. Ассоциативные нейроны находятся в боковых рогах спинного мозга. Здесь нервные импульсы переключаются на промежуточные преганглионарные нейроны, отростки которых покидают центральные ядра и достигают вегетативных ганглиев, где передают импульсы на двигательный нейрон. В связи с этим различают нервные волокна преганглионарные и постганглионарные. Первые из них покидают центральную нервную систему в составе вентральных корешков спинномозговых нервов и черепных нервов. Как в симпатической, так и в парасимпатической системах преганглионарные нервные волокна принадлежат холинергическим нейронам. Аксоны нейронов, расположенных в вегетативных ганглиях, называются постганглионарными. Они не образуют прямых контактов с эффекторными клетками. Их терминальные отделы по своему ходу формируют расширения - варикозности, в составе которых находяся пузырьки медиатора. В области варикозности нет глиальной оболочки и нейромедиатор, выделяясь в окружающую среду, влияет на эффекторные клетки (например, на клетки желез, гладкие миоциты и др.).
В периферических ганглиях симпатической нервной системы, как правило, находятся адренергические эфферентные нейроны (за исключением нейронов, имеющих синаптические связи с потовыми железами, где симпатические нейроны являются холинергическими). В парасимпатических ганглиях эфферентные нейроны всегда холинергические.
Ганглии представляют собой скопления мультиполярных нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия. Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны. В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля I-го типа), чувствительные (клетки Догеля II-го типа) и ассоциативные (клетки Догеля III-го типа). Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки I-го типа называют длинноаксонными нейронами. Нейроны II-го типа - равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки II-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля III-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
Учебное видео анатомии вегетативной нервной системы (ВНС)
При проблемах с просмотром скачайте видео со страницыМышечные ткани - это ткани, отличающиеся по структуре и происхождению, но имеют общую способность к сокращению. Состоят из миоцитов - клеток, которые могут воспринимать нервные импульсы и отвечать на них сокращением.
Свойства и виды мышечной ткани
Морфологические признаки:
- Вытянутая форма миоцитов;
- продольно размещены миофибриллы и миофиламенты;
- митохондрии находятся вблизи сократительных элементов;
- присутствуют полисахариды, липиды и миоглобин.
Свойства мышечной ткани:
- Сократимость;
- возбудимость;
- проводимость;
- растяжимость;
- эластичность.
Выделяют следующие виды мышечной ткани в зависимости от морфофункциональных особенностей:
- Поперечнополосатая: скелетная, сердечная.
- Гладкая.
Гистогенетическая классификация делит мышечные ткани на пять видов в зависимости от эмбрионального источника:
- Мезенхимные - десмальный зачаток;
- эпидермальные - кожная эктодерма;
- нейральные - нервная пластинка;
- целомические - спланхнотомы;
- соматические - миотом.
Из 1-3 видов развиваются гладкомышечные ткани, 4, 5 дают поперечнополосатые мышцы.
Строение и функции гладкой мышечной ткани
Cостоит из отдельных мелких веретеновидных клеток. Эти клетки имеют одно ядро и тонкие миофибриллы, которые тянутся от одного конца клетки к другому. Гладкие мышечные клетки объединяются в пучки, состоящие из 10-12 клеток. Это объединение возникает благодаря особенностям иннервации гладкой мускулатуры и облегчает прохождение нервного импульса на всю группу гладких мышечных клеток. Сокращается гладкая мышечная ткань ритмично, медленно и на протяжении длительного времени, способна при этом развивать большую силу без значительных затрат энергии и без утомления.
У низших многоклеточных животных из гладкой мышечной ткани состоят все мышцы, тогда как у позвоночных животных она входит в состав внутренних органов (кроме сердца).
Сокращения этих мышц не зависят от воли человека, т. е. происходят непроизвольно.
Функции гладкой мышечной ткани:
- Поддерживание стабильного давления в полых органах;
- регуляция уровня кровяного давления;
- перистальтика пищеварительного тракта, перемещения по нему содержимого;
- опорожнение мочевого пузыря.
Строение и функции скелетной мышечной ткани

Cостоит из длинных и толстых волокон длиной 10-12 см. Скелетная мускулатура характеризуется произвольным сокращением (в ответ на импульсы, идущие из коры головного мозга). Скорость ее сокращения в 10-25 раз выше, чем в гладкой мышечной ткани.
Мышечное волокно поперечнополосатой ткани покрыто оболочкой - сарколеммой. Под оболочкой находится цитоплазма с большим количеством ядер, расположенных по периферии цитоплазмы, и сократительными нитями - миофибриллами. Состоит миофибрилла из последовательно чередующихся темных и светлых участков (дисков), обладающих разным коэффициентом преломления света. С помощью электронного микроскопа установлено, что миофибрилла состоит из протофибрилл. Тонкие протофибриллы построены из белка - актина, аболее толстые - из миозина.
При сокращении волокон происходит возбуждение сократимых белков, тонкие протофибриллы скользят по толстым. Актин реагирует с миозином, и возникает единая актомиозиновая система.
Функции скелетной мышечной ткани:
- Динамическая - перемещение в пространстве;
- статическая - поддержание определенной позиции частей тела;
- рецепторная - проприорецепторы, воспринимающие раздражение;
- депонирующая - жидкость, минералы, кислород, питательные вещества;
- терморегуляция - расслабление мышц при повышении температуры для расширения сосудов;
- мимика - для передачи эмоций.
Строение и функции сердечной мышечной ткани
 Сердечная мышечная ткань
Сердечная мышечная ткань
Миокард построен из сердечной мышечной и соединительной ткани, с сосудами и нервами. Мышечная ткань относится к поперечнополосатой мускулатуре, исчерченность которой также обусловлена наличием разных типов миофиламентов. Миокард состоит из волокон, которые связаны между собой и формируют сетку. Эти волокна включают одно или двухъядерные клетки, что расположены в виде цепочки. Они получили название сократительных кардиомиоцитов.
Сократительные кардиомиоциты длиной от 50 до 120 микрометров, шириной - до 20 мкм. Ядро здесь располагается в центре цитоплазмы, в отличие от ядер поперечно полосатых волокон. Кардиомиоциты имеют больше саркоплазма и меньше миофибрилл, в сравнении со скелетными мышцами. В клетках сердечной мышцы находится много митохондрий, так как непрерывные сердечные сокращения требуют много энергии.
Вторая разновидность клеток миокарда - это проводящие кардиомиоциты, которые формируют проводящую систему сердца. Проводящие миоциты обеспечивают передачу импульса к сократительным мышечным клеткам.
Функции сердечной мышечной ткани:
- Насосная;
- обеспечивает ток крови в кровеносном русле.
Компоненты сократительной системы
Особенности строения мышечной ткани обусловлены выполняемыми функциями, возможностью принимать и проводить импульсы, способностью к сокращению. Механизм сокращения заключается в согласованной работе ряда элементов: миофибрилл, сократительных белков, митохондрий, миоглобина.
В цитоплазме мышечных клеток имеются особые сократительные нити - миофибриллы, сокращение которых возможно при содружественной работе белков - актина и миозина, а также при участии ионов Са. Митохондрии снабжают все процессы энергией. Также энергетические запасы образуют гликоген и липиды. Миоглобин необходим для связывания O 2 и формирование его запаса на период сокращения мышцы, так как во время сокращения идет сдавление кровеносных сосудов и снабжение мышц O 2 резко снижается.
Таблица. Соответствие между характеристикой мышечной ткани и ее видом
| Вид ткани | Характеристика |
|---|---|
| Гладкомышечная | Входит в состав стенок кровеносных сосудов |
| Структурная единица – гладкий миоцит | |
| Сокращается медленно, неосознанно | |
| Поперечная исчерченность отсутствует | |
| Скелетная | Структурная единица – многоядерное мышечное волокно |
| Свойственна поперечная исчерченность | |
| Сокращается быстро, осознанно |
Где находится мышечная ткань?
Гладкие мышцы являются составной частью стенок внутренних органов: желудочно-кишечного тракта, мочеполовой системы, сосудов. Входят в состав капсулы селезенки, кожных покровов, сфинктера зрачка.
Скелетная мускулатуразанимают около 40% от массы тела человека, с помощью сухожилий крепятся к костям. Из этой ткани состоят скелетные мышцы, мышцы рта, языка, глотки, гортани, верхнего участка пищевода, диафрагмы, мимическая мускулатура. Также поперечно полосатые мышцы находится в миокарде.
Чем мышечное волокно скелетной мышцы отличается от гладкой мышечной ткани?
Волокна поперечнополосатых мышц намного длиннее (до 12см), чем клеточные элементы гладкомышечной ткани (0,05-0,4мм). Также скелетные волокна имеют поперечную исчерченность благодаря особому расположению нитей актина и миозина. Для гладких мышц это не характерно.
В мышечных волокнах находится много ядер, а сокращение волокон сильное, быстрое и осознанное. В отличие от гладких мышц, клетки гладкомышечной ткани одноядерные, способны сокращаться в медленном темпе и неосознанно.