Muskuļu un muskuļu kontrakciju bioķīmija. Muskuļu bioķīmija Muskuļu kontrakcijas bioķīmija īsi
Svarīga loma šajā procesā ir kalcija joniem un sarkoplazmas proteīniem – kalsequestrīnam un proteīnam ar augstu afinitāti pret kalciju. Sarkoplazmatiskā retikuluma membrānas ieskauj muskuļu pavedienus. Šie proteīni atrodas SPR cisternās uz iekšējās membrānas, kur saistās Ca 2+ joni. Kalsequestrīns - skābs glikoproteīns (MM 45 000 Da), kas spēj piesaistīt 45 Ca 2+ jonus, proteīns ar augstu afinitāti pret kalciju (MM 55 000 Da) saista 25 Ca 2+ jonus. Ca 2+ pārnešana no tvertnēm notiek pa koncentrācijas gradientu ar vienkāršu difūziju; Ca 2+ pārnešana no citoplazmas uz cisternām notiek pret gradientu, kurā piedalās Ca 2+ atkarīgā ATPāze un ATP. Miera stāvoklī aktīvā transporta sistēma uzglabā kalciju cisternās. Muskuļu kontrakcija sākas ar ierašanos darbības potenciāls uz motora nerva gala plāksnes. Acetilholīns izdalās sinapsē, kas saistās ar muskuļu šķiedras postsinaptiskajiem receptoriem. Turklāt darbības potenciāls izplatās gar sarkolemmu uz T-sistēmas šķērsvirziena kanāliņiem. Z-līniju reģionā signāls tiek pārraidīts no šķērsvirziena kanāliņiem uz sarkoplazmatiskā retikuluma cisternām.
Cisternu membrānu depolarizācija izraisa kalcija izdalīšanos un muskuļu kontrakcijas sākšanos. Kalcijs saistās ar troponīna C apakšvienību. Tas maina visas troponīna molekulas konformāciju – I apakšvienība pārstāj traucēt aktīna mijiedarbību ar miozīnu; T apakšvienības konformācijas izmaiņas tiek pārnestas uz tropomiozīnu. Pēc tam tropomiozīns pagriežas par 20° un atver aktīnā iepriekš slēgtus centrus saistīšanai ar miozīnu. Miozīna galva, kas miera stāvoklī ir ADP + P un + miozīna komplekss, pieķeras aktīnam perpendikulāri, un aktīnam ir augsta afinitāte pret šo kompleksu (šķērsvirziena tiltu veidošanās). Aktīna piesaiste izraisa ātru ADP un Pn izdalīšanos no miozīna. Tas noved pie konformācijas izmaiņām, un miozīna galva griežas par 45 ° (gājiens). Ar aktīnu saistītās galvas rotācija izraisa plāna pavediena pārvietošanos attiecībā pret miozīnu. ATP tiek atkārtoti pievienots miozīna galvai, nevis ADP un Fn, veidojot M + ATP kompleksu. Aktīnam ir zema afinitāte pret to, kas izraisa miozīna galvas atdalīšanu (šķērsvirziena tiltu plīsumu). Tas atkal kļūst perpendikulārs plānajam pavedienam. Miozīna galvā, kas nav saistīta ar aktīnu, notiek ATP hidrolīze. Atkal veidojas ADP + P n + miozīna komplekss, un viss atkārtojas. ATP pievienošana miozīnam un ATP hidrolīze notiek ļoti ātri, bet ADP un Pn hidrolīzes produkti no miozīna tiek atdalīti lēni.
Pēc motora impulsa darbības pārtraukšanas Ca 2+ ar Ca 2+ atkarīgās ATPāzes palīdzību nonāk sarkoplazmatiskajā retikulumā. Kalcija atstāšana no troponīna kompleksa noved pie tropomiozīna pārvietošanās un aktīna aktīvo centru slēgšanas, padarot to nespējīgu mijiedarboties ar miozīnu – muskuļi atslābinās.
Svarīgi muskuļu veselībai neiromuskulārās transmisijas procesi. viens) Ar myasthenia gravis asinīs tiek konstatētas antivielas pret saviem acetilholīna receptoriem, kas izpaužas kā muskuļu vājums.
2).
Vairākas zāles (atropīns, sukcinilholīns, kurāra inde) inhibē receptoru proteīni, h Tas bloķē neiromuskulāro vadīšanu.
3).
Narkotikas (neostigmīns, ezerīns) kavē acetilholīna esterāze tādējādi pastiprinot acetilholīna darbību.
4).
Spēcīgāki enzīmu inhibitori ir organiskie fluorofosfāti. Tie veido spēcīgu saikni ar acetilholīnesterāzi un izraisa nāvi no elpošanas apstāšanās. Tās ir nervu indes – tabuns, zarīns.
Cikliskās bioķīmiskās reakcijas, kas notiek muskulī kontrakcijas laikā, nodrošina atkārtotu saaugumu veidošanos un iznīcināšanu starp "galvām" - biezu protofibrilu miozīna molekulu izaugumiem un izvirzījumiem - tievo protofibrilu aktīviem centriem. Darbs pie adhēziju veidošanās un aktīna pavediena veicināšanas gar miozīna pavedienu prasa gan precīzu kontroli, gan ievērojamus enerģijas izdevumus. Reāli šķiedru saraušanās brīdī katrā aktīvajā centrā – dzega – veidojas aptuveni 300 saaugumi minūtē.
Kā jau minēts iepriekš, tikai ATP enerģiju var tieši pārvērst muskuļu kontrakcijas mehāniskajā darbā. ATP, ko hidrolizē miozīna enzīmu centrs, veido kompleksu ar visu proteīna miozīnu. ATP-miozīna kompleksā, kas piesātināts ar enerģiju, miozīns maina savu struktūru un līdz ar to ārējos “izmērus” un tādā veidā veic mehānisku darbu, lai saīsinātu miozīna pavediena izaugumu.
Atpūtas muskuļos miozīns joprojām ir saistīts ar ATP, bet caur Mg ++ joniem bez hidrolītiskas ATP šķelšanās. Miozīna un aktīna saķeres veidošanos miera stāvoklī novērš tropomiozīna komplekss ar troponīnu, kas bloķē aktīna aktīvos centrus. Blokāde tiek saglabāta un ATP netiek sadalīts, kamēr Ca ++ joni ir saistīti. Kad nervu impulss nonāk pie muskuļu šķiedras, tas atbrīvo impulsa raidītājs- neirohormons acetilholīns. Ar Na + joniem negatīvais lādiņš uz sarkolemmas iekšējās virsmas tiek neitralizēts un notiek tā depolarizācija. Šajā gadījumā Ca ++ joni tiek atbrīvoti un saistās ar troponīnu. Savukārt troponīns zaudē lādiņu, kādēļ tiek atbrīvoti aktīvie centri - aktīna pavedienu izvirzījumi un starp aktīnu un miozīnu parādās saaugumi (jo tievo un biezo protofibrilu elektrostatiskā atgrūšana jau ir noņemta). Tagad Ca ++ klātbūtnē ATP mijiedarbojas ar miozīna fermentatīvās aktivitātes centru un sadalās, un pārveidotā kompleksa enerģija tiek izmantota, lai samazinātu saķeres. Iepriekš aprakstītā molekulāro notikumu ķēde ir līdzīga elektriskajai strāvai, kas uzlādē mikrokondensatoru, tās elektriskā enerģija uzreiz tiek pārvērsta mehāniskā darbā uz vietas un ir nepieciešams atkārtoti uzlādēt (ja vēlaties doties tālāk).
Pēc adhēzijas pārrāvuma ATP nesadalās, bet atkal veido enzīmu-substrāta kompleksu ar miozīnu:
M–A + ATP -----> M – ATP + A vai
M-ADP-A + ATP ----> M-ATP + A + ADP
Ja šajā brīdī pienāk jauns nervu impulss, tad “uzlādes” reakcijas atkārtojas, ja nākamais impulss nepienāk, muskulis atslābst. Savilktā muskuļa atgriešanos relaksācijas laikā sākotnējā stāvoklī nodrošina muskuļu stromas proteīnu elastīgie spēki. Izvirzot mūsdienu hipotēzes par muskuļu kontrakciju, zinātnieki ierosina, ka kontrakcijas brīdī aktīna pavedieni slīd pa miozīna pavedieniem, un to saīsināšana iespējama arī kontraktilo proteīnu telpiskās struktūras izmaiņu dēļ (spirāles formas izmaiņas).
Miera stāvoklī ATP ir plastificējoša iedarbība: savienojoties ar miozīnu, tas novērš tā saķeres veidošanos ar aktīnu. Sadaloties muskuļu kontrakcijas laikā, ATP nodrošina enerģiju adhēzijas saīsināšanas procesam, kā arī "kalcija sūkņa" darbam - Ca ++ jonu padevei.ATP šķelšanās muskulī notiek ļoti lielā ātrumā: uz augšu līdz 10 mikromoliem uz 1 g muskuļu minūtē. Tā kā kopējās ATP rezerves muskuļos ir nelielas (ar tām var pietikt tikai 0,5-1 sek darbam ar maksimālo jaudu), lai nodrošinātu normālu muskuļu darbību, ATP ir jāatjauno tādā pašā ātrumā, kādā tas tiek sadalīts.
Lekcija Nr.4. Enerģija muskuļu kontrakcijai, muskuļu darba laikā notiekošie bioķīmiskie procesi.
Glābšanas resintēze.
Konkrēti, tikai ATP var pārveidot ķīmisko enerģiju (tās brīvo daļu, kas atrodas fosfātu saitēs) kustības mehāniskajā enerģijā (lidojums, skriešana un slīdēšana). Viņa ir nodrošina enerģiju attiecīgi adhēzijas saīsināšanas process, vispārēja muskuļu kontrakcija un arī piegādā enerģiju kontrakcijā iesaistīto Ca ++ jonu veidošanai). Dzīva šūna pastāvīgi uztur ATP darba koncentrāciju aptuveni 0,25% līmenī, tostarp intensīva muskuļu darba laikā. Ja (vielmaiņas traucējumu gadījumā) palielinās ATP koncentrācija, tad tiks traucēta muskuļa kontraktilitāte (tā izskatīsies pēc “lupatas”), ja samazināsies, rodas stīvums - stāvoklis pastāvīga nepārejoša kontrakcija ("pārakmeņošanās"). ATP darba koncentrācija ir pietiekama sekundei spēcīgam darbam (3-4 atsevišķas kontrakcijas). Ilgstošas muskuļu darbības laikā ATP darba koncentrācija tiek uzturēta, pateicoties reakcijām tās atjaunošanai. Lai vielmaiņas procesā nodrošinātu normālu (ilgstošu) muskuļu darbu, ATP tiek atjaunots tādā pašā ātrumā, kādā tas tiek sadalīts.
Atgādiniet, ka ATP sadalīšanās ir fermentatīvā hidrolīzes reakcija, un to var izteikt ar vienādojumu:
ATP-āze + ATP + H2O ---> ADP + H3PO4
Enerģija ATP resintēzei (tā pēc tam izdalīsies sadalīšanas laikā - apmēram 40 kJ uz 1 mol) jāiegūst reakcijās, kas notiek ar enerģijas izdalīšanos (kataboliskā). Tāpēc šūnu līmenī ATP hidrolīzes reakcija ir saistīta ar reakcijām, kas nodrošina ATP resintēzi. Šādu reakciju gaitā veidojas vidēji augstas enerģijas savienojumi, kuru sastāvā ir fosfātu grupa, kas kopā ar brīvās enerģijas rezervi tiek pārnesta uz ADP. Šādas pārnešanas reakcijas ("stafetes" pārneses), ko katalizē fosfotransferāzes enzīmi, sauc par transfosforilēšanas vai refosforilēšanas reakcijām. ATP resintēzei nepieciešamie makroerģiskie savienojumi vai nu pastāvīgi atrodas, piemēram, kreatīna fosfāts (uzkrājas simpplastā), vai arī veidojas (difosfoglicerīnskābe, fosfopiruvskābe) oksidatīvos procesos (kataboliskā).
ATP resintēzi muskuļu aktivitātes laikā var veikt divos veidos: reakciju rezultātā bez skābekļa līdzdalības - anaerobā (kad skābekļa piegāde muskuļiem nav laika vai ir sarežģīta) un oksidatīvo procesu dēļ šūnās (piedaloties skābeklis, ko mēs elpojam un ko sportists ātri ieelpo zem slodzes un sākuma atpūtas fāzē).
Cilvēka skeleta muskuļos ir identificēti trīs veidu anaerobie procesi, kuru laikā tiek veikta ATP resintēze:
- kreatīna fosfokināzes reakcija (fosfogēns vai laktisks anaerobs process), kur ATP resintēze notiek pārfosforilācijas dēļ starp kreatīna fosfātu un ADP;
- glikolīze (laktacīda anaerobs process), kur ogļhidrātu fermentatīvās anaerobās sadalīšanās gaitā tiek veikta ATP resintēze, kas beidzas ar pienskābes veidošanos.
- miokināzes reakcija, kurā tiek veikta ATP resintēze noteiktas ADP daļas defosforilēšanas dēļ;
Lai salīdzinātu un kvantitatīvi noteiktu dažāda veida enerģijas konversijas procesus muskuļu aktivitātes laikā, tiek izmantoti trīs galvenie kritēriji:
- jaudas kritērijs - norāda enerģijas konversijas ātrumu noteiktā procesā (vingrinājumā);
- jaudas kritērijs - atspoguļo kopējās enerģētisko vielu rezerves (mēra pēc izdalītās enerģijas daudzuma un veiktā darba);
- efektivitātes kritērijs - raksturo attiecību starp ATP resintēzei iztērēto enerģiju un kopējo šī procesa (vingrinājuma) laikā izdalītās enerģijas daudzumu.
Enerģijas pārveidošanas procesi, anaerobie un aerobie, atšķiras pēc jaudas, jaudas un efektivitātes. Anaerobie procesi dominē īslaicīgos augstas intensitātes vingrinājumos, aerobos - ilgstošas vidējas intensitātes darba laikā.
Lekcijas kopsavilkums| Lekcijas kopsavilkums | Interaktīvs tests | Lejupielādēt abstraktu
» Skeleta muskuļu strukturālā organizācija
» Skeleta muskuļu kontrakcijas molekulārie mehānismi
» Uzbudinājuma un kontrakcijas savienošana skeleta muskuļos
» Skeleta muskuļu relaksācija
»
» Skeleta muskuļu darbs
» Gludo muskuļu strukturālā organizācija un kontrakcijas
» Muskuļu fizioloģiskās īpašības
Muskuļu kontrakcija ir svarīga ķermeņa funkcija, kas saistīta ar aizsardzības, elpošanas, uztura, seksuālajiem, ekskrēcijas un citiem fizioloģiskiem procesiem. Visu veidu brīvprātīgas kustības – staigāšanu, sejas izteiksmes, acs ābolu kustības, rīšanu, elpošanu u.c. veic skeleta muskuļi. Patvaļīgas kustības (izņemot sirds kontrakciju) - kuņģa un zarnu peristaltiku, asinsvadu tonusa izmaiņas, urīnpūšļa tonusa saglabāšanu - izraisa gludo muskuļu kontrakcija. Sirds darbu nodrošina sirds muskuļu kontrakcija.
Skeleta muskuļu strukturālā organizācija
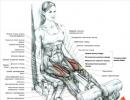
Muskuļu šķiedra un miofibrila (1. att.). Skeleta muskuļi sastāv no daudzām muskuļu šķiedrām, kurām ir stiprinājuma punkti pie kauliem un kas ir paralēli viens otram. Katra muskuļu šķiedra (miocīts) ietver daudzas apakšvienības – miofibrilus, kas veidotas no gareniski atkārtojošiem blokiem (sarkomēriem). Sarkomērs ir skeleta muskuļu saraušanās aparāta funkcionālā vienība. Miofibrils muskuļu šķiedrās atrodas tā, ka sarkomēru atrašanās vieta tajās sakrīt. Tādējādi tiek izveidots šķērsvirziena svītrojums.
Sarkomērs un pavedieni. Sarkomēri miofibrilā ir atdalīti viens no otra ar Z-plāksnēm, kas satur proteīnu beta-aktinīns. Abos virzienos no Z veida plāksnes stiepjas plāni aktīna pavedieni. Starp tiem ir biezāki miozīna pavedieni.
Aktīna pavediens izskatās kā divas lodītes, kas savītas dubultā spirālē, kur katra lodīte ir aktīna proteīna molekula. Aktīna spirāļu padziļinājumos vienādā attālumā viena no otras atrodas troponīna proteīna molekulas, kas savienotas ar pavedienveida tropomiozīna proteīna molekulām.
Miozīna pavedienus veido atkārtotas miozīna proteīna molekulas. Katrai miozīna molekulai ir galva un aste. Miozīna galva var saistīties ar aktīna molekulu, veidojot tā saukto krusta tiltu.
Muskuļu šķiedras šūnu membrāna veido invaginācijas (šķērsvirziena kanāliņus), kas veic ierosmes vadīšanas funkciju uz sarkoplazmatiskā retikuluma membrānu. Sarkoplazmatiskais tīkls (garenvirziena kanāliņi) ir intracelulārs slēgtu kanāliņu tīkls un veic Ca ++ jonu nogulsnēšanas funkciju.
motora vienība. Skeleta muskuļu funkcionālā vienība ir motora vienība (MU). DE - muskuļu šķiedru kopums, ko inervē viena motora neirona procesi. Vienu MU veidojošo šķiedru ierosme un kontrakcija notiek vienlaicīgi (kad tiek ierosināts attiecīgais motoriskais neirons). Atsevišķi MU var izšaut un sarauties neatkarīgi viens no otra.
Molekulārie kontrakcijas mehānismiskeleta muskulis
Saskaņā ar kvēldiega slīdēšanas teoriju muskuļu kontrakcija notiek aktīna un miozīna pavedienu slīdēšanas dēļ viens pret otru. Vītnes bīdīšanas mehānisms ietver vairākus secīgus notikumus.
Miozīna galviņas pievienojas aktīna pavedienu saistīšanās vietām (2. att., A).
Miozīna mijiedarbība ar aktīnu izraisa miozīna molekulas konformācijas pārkārtošanos. Galvas iegūst ATPāzes aktivitāti un griežas par 120°. Galvu rotācijas dēļ aktīna un miozīna pavedieni viens pret otru pārvietojas "vienu soli" (2.b att.).
Aktīna un miozīna disociācija un galvas konformācijas atjaunošana notiek ATP molekulas pievienošanās miozīna galvai un tās hidrolīzes rezultātā Ca++ klātbūtnē (2. att., C).
Cikls "saistīšanās - konformācijas maiņa - atvienošana - konformācijas atjaunošana" notiek daudzkārt, kā rezultātā aktīna un miozīna pavedieni tiek pārvietoti viens pret otru, sarkomēru Z veida diski tuvojas viens otram un miofibrila saīsinās (att. 2, D).
Uzbudinājuma un kontrakcijas konjugācijaskeleta muskuļos
Miera stāvoklī kvēldiega slīdēšana miofibrilā nenotiek, jo saistīšanās centrus uz aktīna virsmas noslēdz tropomiozīna proteīna molekulas (3. att., A, B). Miofibrilu ierosināšana (depolarizācija) un pareiza muskuļu kontrakcija ir saistīta ar elektromehāniskās savienošanas procesu, kas ietver vairākus secīgus notikumus.
Neiromuskulārās sinapses rezultātā uz postsinaptisko membrānu rodas EPSP, kas rada darbības potenciāla attīstību apgabalā, kas ieskauj postsinaptisko membrānu.
Uzbudinājums (darbības potenciāls) izplatās pa miofibrilu membrānu un sasniedz sarkoplazmas tīklu, pateicoties šķērsenisko kanāliņu sistēmai. Sarkoplazmatiskās retikuluma membrānas depolarizācija noved pie Ca++ kanālu atvēršanās tajā, pa kuriem Ca++ joni nonāk sarkoplazmā (3. att., C).
Ca++ joni saistās ar troponīna proteīnu. Troponīns maina savu konformāciju un izspiež tropomiozīna proteīna molekulas, kas slēdza aktīna saistošos centrus (3.d att.).
Atvērtajiem saistīšanās centriem pievienojas miozīna galviņas, un sākas kontrakcijas process (3. att., E).
Šo procesu attīstībai ir nepieciešams noteikts laika periods (10–20 ms). Laiku no muskuļu šķiedras (muskuļa) ierosināšanas brīža līdz tās kontrakcijas sākumam sauc par latento kontrakcijas periodu.
Skeleta muskuļu relaksācija
Muskuļu relaksāciju izraisa apgrieztā Ca++ jonu pārnešana caur kalcija sūkni sarkoplazmatiskā tīkla kanālos. Tā kā Ca++ tiek izņemts no citoplazmas, paliek arvien mazāk atvērtu saistīšanās vietu, un galu galā aktīna un miozīna pavedieni tiek pilnībā atvienoti; notiek muskuļu relaksācija.
Kontraktūra ir ilgstoša un ilgstoša muskuļu kontrakcija, kas saglabājas pēc stimula pārtraukšanas. Īslaicīga kontraktūra var attīstīties pēc tetāniskas kontrakcijas, kas rodas liela daudzuma Ca ++ uzkrāšanās rezultātā sarkoplazmā; ilgstoša (dažreiz neatgriezeniska) kontraktūra var rasties saindēšanās, vielmaiņas traucējumu rezultātā.
Skeleta muskuļu kontrakcijas fāzes un veidi
Muskuļu kontrakcijas fāzes
Kad skeleta muskuli tiek stimulēts ar vienu virssliekšņa spēka elektriskās strāvas impulsu, notiek viena muskuļa kontrakcija, kurā izšķir 3 fāzes (4. att., A):
latentais (slēptais) kontrakcijas periods (ap 10 ms), kura laikā attīstās darbības potenciāls un notiek elektromehāniskās savienošanās procesi; muskuļu uzbudināmība vienas kontrakcijas laikā mainās atbilstoši darbības potenciāla fāzēm;
saīsināšanas fāze (apmēram 50 ms);
relaksācijas fāze (apmēram 50 ms).
Muskuļu kontrakcijas veidi
Dabiskos apstākļos ķermenī netiek novērota viena muskuļa kontrakcija, jo virkne darbības potenciālu iet gar motoriem nerviem, kas inervē muskuļus. Atkarībā no nervu impulsu biežuma, kas nonāk muskulī, muskulis var sarauties vienā no trim režīmiem (4.b att.).
Atsevišķas muskuļu kontrakcijas notiek ar zemu elektrisko impulsu frekvenci. Ja nākamais impulss nāk muskulim pēc relaksācijas fāzes pabeigšanas, notiek virkne secīgu atsevišķu kontrakciju.
Pie lielākas impulsu frekvences nākamais impulss var sakrist ar iepriekšējā kontrakcijas cikla relaksācijas fāzi. Kontrakciju amplitūda tiks summēta, būs robains stingumkramps - ilgstoša kontrakcija, ko pārtrauc muskuļa nepilnīgas atslābuma periodi.
Turpinot palielināt impulsu biežumu, katrs nākamais impulss iedarbosies uz muskuļu saīsināšanas fāzē, kā rezultātā veidojas gluds stingumkrampji – ilgstoša kontrakcija, ko nepārtrauc relaksācijas periodi.
Frekvences optimālais un pessimums
Tetāniskās kontrakcijas amplitūda ir atkarīga no muskuļu kairinošo impulsu biežuma. Frekvences optimums ir tāda kairinošu impulsu frekvence, pie kuras katrs nākamais impulss sakrīt ar paaugstinātas uzbudināmības fāzi (4. att., A) un attiecīgi izraisa vislielākās amplitūdas stingumkrampjus. Frekvences pessimums ir augstāka stimulācijas frekvence, pie kuras katrs nākamais strāvas impulss nonāk ugunsizturības fāzē (4. att., A), kā rezultātā ievērojami samazinās stingumkrampju amplitūda.
Skeleta muskuļu darbs
Skeleta muskuļu kontrakcijas spēku nosaka 2 faktori:
samazināšanā iesaistīto MU skaits;
muskuļu šķiedru kontrakcijas biežums.
Skeleta muskuļu darbs tiek veikts, koordinēti mainot muskuļu tonusu (spriedzi) un garumu kontrakcijas laikā.
Skeleta muskuļu darba veidi:
dinamisks pārvarēšanas darbs tiek veikts, kad muskulis, saraujoties, kustina ķermeni vai tā daļas telpā;
statisks (turēšanas) darbs tiek veikts, ja muskuļu kontrakcijas dēļ ķermeņa daļas tiek turētas noteiktā stāvoklī;
dinamisks zemāks darbs tiek veikts, ja muskulis funkcionē, bet tajā pašā laikā stiepjas, jo ar spēku, ko tas rada, nepietiek, lai pārvietotu vai noturētu ķermeņa daļas.
Darba laikā muskuļi var sarauties:
izotonisks - muskulis saīsinās ar pastāvīgu sasprindzinājumu (ārēja slodze); izotoniskā kontrakcija tiek reproducēta tikai eksperimentā;
izometrisks - muskuļu sasprindzinājums palielinās, bet tā garums nemainās; muskulis izometriski saraujas, veicot statisku darbu;
auksotonisks - muskuļu sasprindzinājums mainās, saīsinot; auksotoniskā kontrakcija tiek veikta dinamiska pārvarēšanas darba laikā.
Vidējās slodzes noteikums- muskulis var veikt maksimālu darbu ar mērenām slodzēm.
Nogurums ir muskuļa fizioloģisks stāvoklis, kas veidojas pēc ilgstoša darba un izpaužas kā kontrakciju amplitūdas samazināšanās, kontrakcijas latentā perioda pagarināšanās un relaksācijas fāze. Noguruma cēloņi ir: ATP izsīkums, vielmaiņas produktu uzkrāšanās muskuļos. Muskuļu nogurums ritmiskā darba laikā ir mazāks nekā sinapses nogurums. Tāpēc, ķermenim veicot muskuļu darbu, nogurums sākotnēji veidojas CNS sinapses un neiromuskulārās sinapses līmenī.
Strukturālā organizācija un samazināšanagludie muskuļi
Strukturālā organizācija. Gludie muskuļi sastāv no atsevišķām vārpstveida šūnām (miocītiem), kas muskuļos atrodas vairāk vai mazāk nejauši. Kontrakcijas pavedieni ir izkārtoti neregulāri, kā rezultātā nav muskuļa šķērssvītrojuma.
Kontrakcijas mehānisms ir līdzīgs skeleta muskuļos, taču pavedienu slīdēšanas ātrums un ATP hidrolīzes ātrums ir 100–1000 reižu zemāks nekā skeleta muskuļos.
Uzbudinājuma un kontrakcijas konjugācijas mehānisms. Kad šūna ir uzbudināta, Ca++ nonāk miocīta citoplazmā ne tikai no sarkoplazmatiskā tīklekļa, bet arī no starpšūnu telpas. Ca++ joni, piedaloties kalmodulīna proteīnam, aktivizē fermentu (miozīna kināzi), kas pārnes fosfātu grupu no ATP uz miozīnu. Fosforilētas miozīna galviņas iegūst spēju pievienoties aktīna pavedieniem.
Gludo muskuļu kontrakcija un relaksācija. Ca ++ jonu izvadīšanas ātrums no sarkoplazmas ir daudz mazāks nekā skeleta muskuļos, kā rezultātā relaksācija notiek ļoti lēni. Gludie muskuļi veic ilgstošas tonizējošas kontrakcijas un lēnas ritmiskas kustības. Pateicoties zemajai ATP hidrolīzes intensitātei, gludie muskuļi ir optimāli pielāgoti ilgstošai kontrakcijai, kas neizraisa nogurumu un lielu enerģijas patēriņu.
Muskuļu fizioloģiskās īpašības
Skeleta un gludo muskuļu kopējās fizioloģiskās īpašības ir uzbudināmība un kontraktilitāte. Skeleta un gludo muskuļu salīdzinošie raksturlielumi ir doti tabulā. 6.1. Sirds muskuļu fizioloģiskās īpašības un īpatnības ir apskatītas sadaļā "Homeostāzes fizioloģiskie mehānismi".
7.1. tabula. Skeleta un gludo muskuļu salīdzinošās īpašības
|
Īpašums |
Skeleta muskuļi |
Gludie muskuļi |
|
Depolarizācijas ātrums |
lēns |
|
|
Ugunsizturīgs periods |
īss |
garš |
|
Samazinājuma būtība |
ātrā fāze |
lēns toniks |
|
Enerģijas izmaksas |
||
|
Plastmasa |
||
|
Automatizācija |
||
|
Vadītspēja |
||
|
inervācija |
somatiskās NS motoneuroni |
autonomās NS postganglioniskie neironi |
|
Veiktās kustības |
patvaļīgi |
piespiedu kārtā |
|
Jutība pret ķīmiskām vielām |
||
|
Spēja sadalīt un atšķirt |
Gludo muskuļu plastiskums izpaužas faktā, ka tie spēj uzturēt nemainīgu tonusu gan saīsinātā, gan izstieptā stāvoklī.
Gludo muskuļu audu vadītspēja izpaužas faktā, ka ierosme izplatās no viena miocīta uz otru, izmantojot specializētus elektriski vadošus kontaktus (savienojumus).
Gludo muskuļu automātisma īpašība izpaužas faktā, ka tas var sarauties bez nervu sistēmas līdzdalības, jo daži miocīti spēj spontāni radīt ritmiski atkārtojošus darbības potenciālus.
Visi ķermeņa muskuļi ir sadalīti gludos un šķērssvītrotos.
Skeleta muskuļu kontrakcijas mehānismi
Svītrotie muskuļi ir sadalīti divos veidos: skeleta muskuļos un miokardā.
Muskuļu šķiedras struktūra
Muskuļu šūnu membrāna, ko sauc par sarkolemmu, ir elektriski uzbudināma un spēj vadīt darbības potenciālu. Šie procesi muskuļu šūnās notiek pēc tāda paša principa kā nervu šūnās. Muskuļu šķiedras miera potenciāls ir aptuveni -90 mV, t.i., zemāks nekā nervu šķiedras potenciāls (-70 mV); kritiskā depolarizācija, kuru sasniedzot rodas darbības potenciāls, ir tāda pati kā nervu šķiedrai. Tādējādi: muskuļu šķiedras uzbudināmība ir nedaudz zemāka nekā nervu šķiedras uzbudināmība, jo muskuļu šūna ir jādepolarizē lielā daudzumā.
Muskuļu šķiedras reakcija uz stimulāciju ir samazināšana, kas padara šūnas saraušanās aparātu - miofibrils. Tie ir pavedieni, kas sastāv no divu veidu pavedieniem: biezi - miozīns, un tievs aktīns. Biezie pavedieni (15 nm diametrā un 1,5 µm garumā) satur tikai vienu proteīnu – miozīnu. Plānie pavedieni (7 nm diametrā un 1 µm garumā) satur trīs veidu proteīnus: aktīnu, tropomiozīnu un troponīnu.
Aktīns ir garš proteīna pavediens, kas sastāv no atsevišķiem lodveida proteīniem, kas savienoti kopā tā, ka visa struktūra ir iegarena ķēde. Globulārā aktīna (G-aktīna) molekulām ir sānu un gala saistīšanās vietas ar citām līdzīgām molekulām. Rezultātā tie ir apvienoti tā, ka veido struktūru, ko bieži salīdzina ar divām kopā savienotām krellēm. Lente, kas veidojas no G-aktīna molekulām, ir savīta spirālē. Šo struktūru sauc par fibrilāro aktīnu (F-aktīnu). Spirāles solis (spoles garums) ir 38 nm; katrā spirāles spolē ir 7 pāri G-aktīna. G-aktīna polimerizācija, tas ir, F-aktīna veidošanās, notiek ATP enerģijas dēļ, un, gluži pretēji, kad F-aktīns tiek iznīcināts, enerģija atbrīvojas.

1. att. Atsevišķu G-aktīna globulu saplūšana F-aktīnā
Gar aktīna pavedienu spirālveida rievām atrodas proteīns tropomiozīns.Katra tropomiozīna virkne, kuras garums ir 41 nm, sastāv no divām identiskām α ķēdēm, kas savītas kopā spirālē ar 7 nm garu vijumu. Divas tropomiozīna molekulas atrodas vienā F-aktīna pagriezienā. Katra tropomiozīna molekula nedaudz pārklājas ar nākamo, kā rezultātā veidojas tropomiozīna pavediens, kas nepārtraukti stiepjas gar aktīnu.

2. att. Plāna miofibrila pavediena struktūra
Svītrotās muskuļu šūnās papildus aktīnam un tropomiozīnam tievu pavedienu sastāvā ietilpst arī proteīns troponīns. Šim lodveida proteīnam ir sarežģīta struktūra. Tas sastāv no trim apakšvienībām, no kurām katra veic savu funkciju kontrakcijas procesā.
biezs pavediens sastāv no daudzām molekulām miozīns savākti saišķī. Katra 155 nm gara un 2 nm diametra miozīna molekula sastāv no sešiem polipeptīdu pavedieniem: diviem gariem un četriem īsiem. Garās ķēdes ir satītas kopā 7,5 nm piķa spirālē un veido miozīna molekulas fibrilāro daļu. Vienā molekulas galā šīs ķēdes atritinās un veido dakšveida galu. Katrs no šiem galiem veido kompleksu ar divām īsām ķēdēm, tas ir, uz katras molekulas ir divas galvas. Šī ir miozīna molekulas lodveida daļa.

3. att. Miozīna molekulas struktūra.
Miozīnā izšķir divus fragmentus: vieglo meromiozīnu (LMM) un smago meromiozīnu (HMM), starp tiem ir eņģe. TMM sastāv no diviem apakšfragmentiem: S1 un S2. LMM un apakšfragments S2 ir ligzdoti pavedienu saišķī, un apakšfragments S1 izvirzīts virs virsmas. Šis izvirzītais gals (miozīna galva) spēj saistīties ar aktīna pavediena aktīvo vietu un mainīt slīpuma leņķi pret miozīna pavedienu saišķi. Atsevišķu miozīna molekulu apvienošana saišķī notiek elektrostatiskās mijiedarbības dēļ starp LMM. Vītnes centrālajai daļai nav galviņu. Viss miozīna molekulu komplekss pārsniedz 1,5 µm. Tā ir viena no lielākajām dabā zināmajām bioloģiskajām molekulārajām struktūrām.
Aplūkojot šķērssvītrota muskuļa garengriezumu caur polarizējošo mikroskopu, ir redzami gaiši un tumši apgabali. Tumšie apgabali (diski) ir anizotropi: polarizētā gaismā tie izskatās caurspīdīgi garenvirzienā un necaurredzami šķērsvirzienā, ko apzīmē ar burtu A. Gaismas zonas ir izotropas un apzīmētas ar burtu I. Disks I ietver tikai plānus pavedienus , un disks A - un biezs un plāns. Diska A vidū ir redzama gaismas josla, ko sauc par H zonu. Tam nav plānu pavedienu. Disks I ir sadalīts ar plānu sloksni Z, kas ir membrāna, kurā ir strukturālie elementi, kas savieno kopā tievu pavedienu galus. Tiek saukts laukums starp divām Z-līnijām sarkomere.

4. att. Miofibrilu struktūra (šķērsgriezums)

5. att. Svītrotā muskuļa struktūra (garengriezums)
Katru resno pavedienu ieskauj seši tievie pavedieni, un katru tievo pavedienu ieskauj trīs biezi. Tādējādi šķērsgriezumā muskuļu šķiedrai ir regulāra sešstūra struktūra.
Muskuļu kontrakcija
Muskuļu kontrakcijas laikā aktīna un miozīna pavedienu garums nemainās. Ir tikai to pārvietošanās viens pret otru: plāni pavedieni pārvietojas spraugā starp biezajiem. Šajā gadījumā diska A garums paliek nemainīgs, un disks I saīsinās, sloksne H gandrīz pazūd. Šāda slīdēšana ir iespējama, jo starp bieziem un plāniem pavedieniem ir šķērseniski tilti (miozīna galvas). Ar kontrakciju ir iespējamas sarkomēra garuma izmaiņas no aptuveni 2,5 līdz 1,7 mikroniem.
Miozīna pavedienam ir daudz galviņu, ar kurām tas var saistīties ar aktīnu. Savukārt aktīna pavedienam ir vietas (aktīvie centri), pie kurām var piestiprināties miozīna galviņas. Atpūtas muskuļu šūnā šīs saistīšanās vietas pārklāj tropomiozīna molekulas, kas novērš saites veidošanos starp plāniem un bieziem pavedieniem.
Lai aktīns un miozīns mijiedarbotos, ir jābūt kalcija joniem. Miera stāvoklī tie atrodas sarkoplazmatiskajā retikulumā. Šī organelle ir membrānas dobums, kurā atrodas kalcija sūknis, kas, izmantojot ATP enerģiju, transportē kalcija jonus sarkoplazmatiskajā retikulumā. Tās iekšējā virsma satur olbaltumvielas, kas spēj saistīt Ca2+, kas nedaudz samazina šo jonu koncentrāciju starpību starp citoplazmu un tīklveida dobumu. Rīcības potenciāls, kas izplatās gar šūnas membrānu, aktivizē tīklveida membrānu, kas atrodas tuvu šūnas virsmai, un izraisa Ca2+ izdalīšanos citoplazmā.
Troponīna molekulai ir augsta afinitāte pret kalciju.
Tās ietekmē tas maina tropomiozīna pavediena stāvokli uz aktīna pavedienu tā, ka atveras aktīvais centrs, ko iepriekš klāja tropomiozīns. Atvērtajam aktīvajam centram pievienojas šķērsenisks tilts. Tas noved pie aktīna mijiedarbības ar miozīnu. Pēc saites veidošanās miozīna galva, kas iepriekš atradās taisnā leņķī pret pavedieniem, sasver un velk aktīna pavedienu attiecībā pret miozīna galvu par aptuveni 10 nm. Izveidotais atīna-miozīna komplekss novērš pavedienu tālāku slīdēšanu vienam pret otru, tāpēc ir nepieciešama tā atdalīšana. Tas ir iespējams tikai ATP enerģijas dēļ. Miozīnam ir ATPāzes aktivitāte, tas ir, tas spēj izraisīt ATP hidrolīzi. Šajā procesā atbrīvotā enerģija sarauj saikni starp aktīnu un miozīnu, un miozīna galva spēj mijiedarboties ar aktīna molekulas jauno daļu. Tiltu darbs tiek sinhronizēts tā, ka visu viena pavediena tiltu sasiešana, sasvēršanās un pārraušana notiek vienlaicīgi. Muskulim atslābinoties, tiek aktivizēts kalcija sūkņa darbs, kas pazemina Ca2+ koncentrāciju citoplazmā; tāpēc vairs nevar veidoties saites starp plāniem un bieziem pavedieniem. Šādos apstākļos, izstiepjot, vītnes muskuļi brīvi slīd viens pret otru. Tomēr šāda paplašināmība ir iespējama tikai ATP klātbūtnē. Ja šūnā nav ATP, tad aktīna-miozīna komplekss nevar salūzt. Vītnes paliek cieši saistītas viena ar otru. Šī parādība tiek novērota rigor mortis.
6. att. Sarkomēra samazināšana: 1 - miozīna pavediens; 2 - aktīvais centrs; 3 - aktīna pavediens; 4 - miozīna galva; 5 - Z-līnija.
a) nav mijiedarbības starp plāniem un bieziem pavedieniem;
b) Ca2+ klātbūtnē miozīna galva saistās ar aktīna pavediena aktīvo vietu;

iekšā)šķērseniskie tilti noliecas un izvelk plānu pavedienu cauri salīdzinoši biezam, kā rezultātā sarkomēra garums samazinās;
G) saites starp pavedieniem tiek pārtrauktas ATP enerģijas ietekmē, miozīna galvas ir gatavas mijiedarboties ar jauniem aktīviem centriem.
Ir divi muskuļu kontrakcijas veidi: izotonisks(šķiedras garums mainās, bet spriegums paliek nemainīgs) un izometrisks(muskuļa gali ir nekustīgi fiksēti, kā rezultātā mainās nevis garums, bet sasprindzinājums).
Muskuļu kontrakcijas spēks un ātrums
Svarīgas muskuļu īpašības ir kontrakciju spēks un ātrums. Vienādojumus, kas izsaka šos raksturlielumus, empīriski ieguva A. Hils, un pēc tam tos apstiprināja muskuļu kontrakcijas kinētiskā teorija (Deščerevska modelis).
Kalna vienādojums, kas attiecas uz muskuļu kontrakcijas spēku un ātrumu, ir šāda forma: (P+a)(v+b) = (P0+a)b = a(vmax+b), kur v ir muskuļu saīsināšanas ātrums; P - muskuļu spēks vai tam pieliktā slodze; vmax ir maksimālais muskuļu saīsināšanas ātrums; P0 ir spēks, ko muskuļi attīsta izometriskā kontrakcijas režīmā; a,b ir konstantes. vispārējā jauda muskuļu attīstīto nosaka pēc formulas: N kopā = (P+a)v = b(P0-P). efektivitāti muskuļi paliek nemainīgi ( apmēram 40%) spēka vērtību diapazonā no 0,2 P0 līdz 0,8 P0. Muskuļu kontrakcijas laikā tiek atbrīvots noteikts siltuma daudzums. Šo vērtību sauc siltuma ražošana. Siltuma ražošana ir atkarīga tikai no muskuļu garuma izmaiņām un nav atkarīga no slodzes. Konstantes a un b ir nemainīgas vērtības konkrētajam muskulim. Pastāvīgi a ir spēka dimensija, un b- ātrums. Pastāvīgi b lielā mērā atkarīgs no temperatūras. Pastāvīgi a ir vērtību diapazonā no 0,25 P0 līdz 0,4 P0. Pamatojoties uz šiem datiem, tiek aprēķināts maksimālais kontrakcijas ātrumsšim muskuļiem: vmax = b (P0/a).
muskuļu audu īpašības.
Skeleta muskuļu kontrakcija un tās mehānismi
Muskuļu audu veidi. Aktīna-miozīna komplekss un tā darbības mehānismi.
Ir 3 dzīvnieku audu veidi: 1) muskuļu, 2) nervu, 3) sekrēcijas. Pirmais reaģē uz ierosmi ar kontrakciju un kustības darba īstenošanu. Otrais - spēja vadīt un analizēt impulsus, trešais - izcelt dažādus noslēpumus.
Ir 3 veidu muskuļu audi: 1. svītraini, 2. gludi, 3. sirds.
| Raksturlielumi | svītraini | gluda | sirds |
| specializācija | ļoti augstu | vismazākā specializācija. | sekundārā specializācija. |
| struktūra | garas šķiedras līdz 10 cm, sadalītas apakšvienībās - sarkomēros. Šķiedras savstarpēji savieno saistaudi, asinsvadi. Nervu gali savienojas ar šķiedrām, veidojot neiromuskulārus savienojumus. | Sastāv no atsevišķas vārpstveida. šūnas, kas savienotas saišķos. | Šūnas galos atzarojas, savienojas viena ar otru ar procesu palīdzību. |
| kodols | Vairāki serdeņi perifērijā | 1 kodols centrā | vairāki serdeņi centrā |
| citoplazma | satur mitohondrijus, sarkoplazmas. tīklojums, T caurules, glikogēns, tauku pilieni | velēna. mitohondriji, sarkoplazma. tīklojums, caurulītes, | velēna. mitohondriji, sarkoplazma. tīklojums, T caurules, |
| sarkolemma | tur ir | Nē | tur ir |
| regulējumu | neirogēns | neirogēns | neirog. un humorāls |
| šķērseniskās svītras | tur ir | Nē | tur ir |
| Savienojuma darbība. | spēcīgas, ātras kontrakcijas. Ugunsizturības periods ir īss - atpūtas laiks ir īss Ātrs nogurums. | lēns ritms | ātrs ritms, ilgs ugunsizturības laiks - nav noguruma. |
Aktīna-miozīna komplekss. Visas muskuļu šūnas satur lielu skaitu īpašu saraušanās proteīnu - tie ir 60-80% no kopējā muskuļu proteīnu daudzuma. galvenais kontraktils
olbaltumvielas ir fibrilāri proteīni: - miozīns- veido biezus pavedienus; — aktīns- veido plānus pavedienus. Kontrakcijas regulēšanai tiek izmantoti lodveida proteīni: troponīns-tropomiozīns.
Miozīns ir 2-šķiedru struktūra 1=180 nm un 0=2,5 nm. Aktīns ir 2-spirāles peptīdu ķēde.
Samazināšanas mehānisms: Aktīns un miozīns fibrilā ir telpiski atdalīti. Nervu impulss izraisa acetilholīna izdalīšanos neiromuskulārā savienojuma sinaptiskajā plaisā. to
izraisa postsinaptiskās membrānas depolarizāciju pēc neirotransmitera saistīšanās un
darbības potenciāla izplatīšanās cauri šūnu membrānām un muskuļos
šķiedras caur T caurulēm. Aktīna-miozīna mijiedarbības rezultātā notiek fibrilu kontrakcija. Tas tiek panākts, tilta veidošanās rezultātā nospiežot aktīna pavedienu ar miozīna galvu. Kad impulss pazūd, Ca2+ tiek atjaunots, tilts starp aktīnu un miozīnu tiek iznīcināts un muskulis atgriežas sākotnējā stāvoklī.
Troponīns ir lodveida proteīns ar 3 centriem:
- T - saistās ar tropomiozīnu
- C - saistās ar Ca2 +
- 1 - kavē aktīna-miozīna mijiedarbību.
Kontrakcijas fāzes:
1. Latentais periods - 0,05 sek.
2. Kontrakcijas fāze - 0,1 sek
3. Relaksācijas periods - 0,2 sek.
Muskuļu darba bioķīmija
1. ATP + miozīna-aktīna komplekss—-ADP + miozīns + aktīns + F + enerģija
2. ADP + kreatinīna fosfāts — ATP + kreatīns
3. Glikogēns-glikoze--glikoze + O2--CO2 + H2O + 38 ATP (aerobs process)
4. Glikoze - 2 pienskābe + 2 ATP
5. Piena skābe + O2-CO2 + H2O (atpūta) vai Mol.k-ta-glikoze-glikogēns.
Skeleta muskuļu kontrakcijas mehānisms
Muskuļu saīsināšana ir vairāku sarkomēru kontrakcijas rezultāts. Saīsinot aktīna pavedienus, tie slīd attiecībā pret miozīna pavedieniem, kā rezultātā samazinās katra muskuļu šķiedras sarkomēra garums. Šajā gadījumā pašu pavedienu garums paliek nemainīgs. Miozīna pavedieniem ir aptuveni 20 nm gari šķērsvirziena izvirzījumi (šķērsojošie tilti). Katrs izvirzījums sastāv no galviņas, kas caur "kaklu" savienota ar miozīna pavedienu (23. att.).
Atvieglinātā stāvoklī šķērsenisko tiltu galvas muskuļi nevar mijiedarboties ar aktīna pavedieniem, jo to aktīvās vietas (savstarpēja kontakta vietas ar galvām) izolē tropomiozīns. Muskuļa saīsināšana ir šķērseniskā tilta konformācijas izmaiņu rezultāts: tā galva noliecas, saliekot “kaklu”.

Rīsi. 23. Kontrakcijas un regulējošo proteīnu telpiskā organizācija šķērssvītrotā muskulī. Miozīna tilta stāvoklis (insulta efekts, kakls ir saliekts) tiek parādīts kontraktilo proteīnu mijiedarbības procesā muskuļu šķiedrās (šķiedru kontrakcija)
Procesa secība , nodrošinot muskuļu šķiedru kontrakcija(elektromehāniskā saskarne):
1. Pēc notikuma PD muskuļšķiedrā pie sinapses (PCP elektriskā lauka dēļ) ierosme izplatās pa miocītu membrānu, ieskaitot šķērsvirziena membrānas T veida kanāliņi. AP vadīšanas mehānisms pa muskuļu šķiedru ir tāds pats kā pa nemielinizētu nervu šķiedru - radusies AP sinapses tuvumā caur tās elektrisko lauku nodrošina jaunu AP rašanos blakus šķiedras sadaļā utt. (nepārtraukta ierosmes vadīšana).
2. Potenciāls darbības T veida kanāliņi tā elektriskā lauka dēļ tiek aktivizēti ar spriegumu saistīti kalcija kanāli membrāna SPR, kā rezultātā Ca2+ atstāj SPR tvertnes saskaņā ar elektroķīmisko gradientu.
3. Starpfibrilārajā telpā Ca2+ kontakti ar troponīns, kas noved pie tā konformācijas un tropomiozīna pārvietošanas, kā rezultātā veidojas aktīna pavedieni tiek atklātas aktīvās zonas ar ko tie ir saistīti miozīna tiltu galvas.
4. Mijiedarbības ar aktīnu rezultātā Tiek pastiprināta miozīna pavedienu galviņu ATPāzes aktivitāte, nodrošinot ATP enerģijas izdalīšanos, kas tiek tērēta miozīna tilta locīšanaārēji atgādina airu kustību airēšanas laikā (airēšanas kustība) (sk. 23. att.), nodrošina aktīna pavedienu slīdēšanu attiecībā pret miozīna pavedieniem. Viena insulta pabeigšanai ir nepieciešama vienas ATP molekulas enerģija. Šajā gadījumā kontraktilo proteīnu virknes tiek pārvietotas par 20 nm. Jaunas ATP molekulas pievienošana citai miozīna galvas daļai noved pie tās iesaistīšanās pārtraukšanas, bet ATP enerģija netiek patērēta. Ja nav ATP, miozīna galviņas nevar atrauties no aktīna – muskulis ir saspringts; jo īpaši tāds ir rigor mortis mehānisms.
5. Pēc tam krustenisko tiltu galviņas to elastības dēļ atgriežas sākotnējā stāvoklī un izveido kontaktu ar nākamo aktīna vietu; tad atkal notiek cita airēšanas kustība un aktīna un miozīna pavedienu slīdēšana. Šādas elementāras darbības tiek atkārtotas daudzas reizes. Viens sitiens (viens solis) izraisa katra sarkomēra garuma samazināšanos par 1%. Ar izolētas vardes muskuļa kontrakciju bez slodzes par 50%, sarkomēru saīsināšana notiek 0,1 s. Tam nepieciešamas 50 airēšanas kustības.
Muskuļu kontrakcijas mehānisms
Miozīna tilti liecas asinhroni, bet, ņemot vērā to, ka to ir daudz un katru miozīna pavedienu ieskauj vairāki aktīna pavedieni, muskuļu kontrakcija notiek vienmērīgi.
Relaksācija muskuļi ir saistīti ar procesiem, kas notiek apgrieztā secībā. Sarkolemmas un T-kanāliņu repolarizācija noved pie SPR membrānas kalcija sprieguma kanālu slēgšanas. Ca-sūkņi atdod Ca2+ SPR (sūkņu aktivitāte palielinās, palielinoties brīvo jonu koncentrācijai).
Ca2+ koncentrācijas samazināšanās starpfibrilārajā telpā izraisa apgrieztu troponīna konformāciju, kā rezultātā tropomiozīna pavedieni izolē aktīna pavedienu aktīvās vietas, kas padara neiespējamu miozīna krustenisko tiltu galvām mijiedarbību ar tiem. Aktīna pavedienu slīdēšana pa miozīna pavedieniem pretējā virzienā notiek gravitācijas spēku un muskuļu šķiedru elementu elastīgās vilces ietekmē, kas atjauno sarkomēru sākotnējos izmērus.
ATP ir enerģijas avots skeleta muskuļu darba nodrošināšanai, kura izmaksas ir ievērojamas. Pat muskuļu funkcionēšanas galvenās apmaiņas apstākļos ķermenis ietekmē apmēram 25% no visiem saviem enerģijas resursiem. Fiziskā darba veikšanas laikā enerģijas izmaksas krasi pieaug.
ATP rezerves muskuļu šķiedrās ir nenozīmīgas (5 mmol / l) un var nodrošināt ne vairāk kā 10 atsevišķas kontrakcijas.
Enerģijas patēriņš ATP ir nepieciešams šādiem procesiem.
Pirmkārt, ATP enerģija tiek tērēta, lai nodrošinātu Na/K-sūkņa darbību (tas uztur Na+ un K+ koncentrācijas gradientu šūnas iekšpusē un ārpusē, kas veido PP un PD, kas nodrošina elektromehānisko savienojumu) un sūkņa darbību. Ca-sūknis, kas pazemina Ca2+ koncentrāciju sarkoplazmā pēc muskuļu šķiedras kontrakcijas, kas noved pie relaksācijas.
Otrkārt, ATP enerģija tiek tērēta miozīna tiltu airēšanas kustībai (to locīšanai).
ATP resintēze tiek veikta ar trīs ķermeņa enerģētisko sistēmu palīdzību.
1. Fosfogēnās enerģijas sistēma nodrošina ATP resintēzi, pateicoties muskuļos esošajam ļoti energoietilpīgajam CP un adenozīna difosfātam (adenozīndifosfāts, ADP), kas veidojas ATP sadalīšanās laikā, veidojoties kreatīnam (K): ADP + + CF → ATP + K. Tā ir tūlītēja ATP resintēze, kamēr muskulis var attīstīt lielu jaudu, bet īsu laiku - līdz 6 s, jo CF rezerves muskuļos ir ierobežotas.
2. Anaerobā glikolītiskā enerģijas sistēma nodrošina ATP resintēzi, pateicoties enerģijai, kas rodas glikozes anaerobajā sadalīšanā pienskābē. Šis ATP resintēzes ceļš ir ātrs, bet arī īslaicīgs (1–2 min), jo pienskābes uzkrāšanās kavē glikolītisko enzīmu aktivitāti. Taču laktāts, izraisot lokālu vazodilatējošo efektu, uzlabo asins plūsmu strādājošajā muskulī un skābekļa un barības vielu piegādi tam.
3. Aerobā enerģijas sistēma nodrošina ATP resintēzi ar palīdzību ogļhidrātu un taukskābju oksidatīvā fosforilēšana kas rodas muskuļu šūnu mitohondrijās. Šādā veidā var nodrošināt enerģiju muskuļu darbam vairākas stundas un ir galvenais veids, kā nodrošināt enerģiju skeleta muskuļu darbam.
Muskuļu kontrakciju veidi
Atkarībā no kontrakciju rakstura Ir trīs veidu muskuļi: izometriski, izotoniski un auksotoniski.
Muskuļa auksotoniskā kontrakcija sastāv no vienlaicīgas muskuļa garuma un spriedzes izmaiņām. Šāda veida kontrakcijas ir raksturīgas dabiskām motora darbībām un var būt divu veidu: ekscentriskas, kad muskuļu sasprindzinājumu pavada tā pagarināšanās - piemēram, tupēšanas (nolaišanas) procesā, un koncentriskas, kad muskuļu sasprindzinājumu pavada tā sasprindzinājums. saīsināšana - piemēram, pagarinot apakšējās ekstremitātes pēc tupēšanas (kāpšanas).
Izometriskā muskuļu kontrakcija- kad palielinās muskuļu sasprindzinājums un tā garums nemainās. Šāda veida kontrakcijas var novērot eksperimentā, kad abi muskuļa gali ir fiksēti un nav iespējama to saplūšana, un dabiskos apstākļos - piemēram, pietupiena un pozīcijas fiksēšanas procesā.
Izotoniskā muskuļu kontrakcija Tas sastāv no muskuļu saīsināšanas ar pastāvīgu spriedzi. Šāda veida kontrakcijas rodas, ja nenoslogots muskulis ar vienu piestiprinātu cīpslu saraujas, nepaceļot (pārvietojot) nekādu ārēju slodzi vai paceļot slodzi bez paātrinājuma.
Atkarībā no ilguma Ir divu veidu muskuļu kontrakcijas: vientuļnieks un tetānisms.
Viena muskuļa kontrakcija rodas ar vienreizēju nerva vai paša muskuļa kairinājumu. Parasti muskulis tiek saīsināts par 5-10% no tā sākotnējā garuma. Uz vienas kontrakcijas līknes ir trīs galvenie periodi: 1) latentais- laiks no kairinājuma pielietošanas brīža līdz kontrakcijas sākumam; 2) periods saīsināšana (vai stresa attīstība); 3) periods relaksācija. Viena cilvēka muskuļu kontrakciju ilgums ir mainīgs. Piemēram, zoles muskulī tas ir 0,1 s. Latentajā periodā notiek muskuļu šķiedru uzbudinājums un tā vadīšana gar membrānu. Muskuļu šķiedras vienas kontrakcijas ilguma, tās ierosmes un fāzes izmaiņu attiecība muskuļu šķiedras uzbudināmībā ir parādīta attēlā. 24.
Muskuļu šķiedru kontrakcijas ilgums ir daudz ilgāks nekā AP, jo ir nepieciešams laiks, lai Ca sūkņi darbotos, lai atgrieztu Ca2+ SPR un vidē, un lielāka mehānisko procesu inerce, salīdzinot ar elektrofizioloģiskajiem.

Rīsi. 24. att. Siltasiņu skeleta muskuļa lēnās šķiedras AP (A) un vienas kontrakcijas (B) rašanās laika attiecība. Bultiņa- kairinājuma brīdis. Ātro šķiedru kontrakcijas laiks ir vairākas reizes īsāks
tetāniskā kontrakcija- tā ir ilgstoša muskuļu kontrakcija, kas notiek ritmiskas stimulācijas ietekmē, kad katrs nākamais kairinājums vai nervu impulss nonāk muskulī, kamēr tas vēl nav atslābināts. Tetāniskā kontrakcija balstās uz atsevišķu muskuļu kontrakciju summēšanas fenomenu (25. att.) – kontrakcijas amplitūdas un ilguma palielināšanos, ja tā tiek pielietota muskuļu šķiedrai vai veselam muskulim no diviem vai vairākiem strauji pēc kairinājumiem.

Rīsi. 25. att. Vardes gastrocnemius muskuļa kontrakciju summēšana: 1 - viena kontrakcijas līkne, reaģējot uz pirmo atslābinātā muskuļa kairinājumu; 2 - viena un tā paša muskuļa vienpusējas kontrakcijas līkne, reaģējot uz otro stimulāciju; 3 - summētās kontrakcijas līkne, kas iegūta saraujošā muskuļa pāra stimulācijas rezultātā ( norādīts ar bultiņām)
Šajā gadījumā kairinājumiem vajadzētu parādīties iepriekšējās kontrakcijas periodā. Kontrakciju amplitūdas palielināšanās ir izskaidrojama ar Ca2+ koncentrācijas palielināšanos hialoplazmā atkārtotas muskuļu šķiedru ierosināšanas laikā, jo Ca sūknim nav laika to atgriezt SPR. Ca2+ nodrošina miozīna tiltu saķeres zonu skaita palielināšanos ar aktīna pavedieniem.
Ja muskuļu relaksācijas fāzē nonāk atkārtoti impulsi vai kairinājumi, tad ir dentate stingumkrampji. Ja atkārtoti kairinājumi iekrīt saīsināšanas fāzē, tas ir gluda stingumkrampji(26. att.).

Rīsi. 26. Vardes gastrocnemius muskuļa kontrakcija pie dažādām sēžas nerva stimulācijas frekvencēm: 1 - vienreizēja kontrakcija (biežums 1 Hz); 2,3 - zobainais stingumkrampji (15-20 Hz); 4,5 - gluda stingumkrampji (25-60 Hz); 6 - relaksācija pie pesimālās stimulācijas frekvences (120 Hz)
Kontrakcijas amplitūda un spriedzes apjoms, ko rada muskuļu šķiedras ar gludu stingumkrampjiem, parasti ir 2-4 reizes lielākas nekā ar vienu kontrakciju. Muskuļu šķiedru stingumiskā kontrakcija, atšķirībā no atsevišķām kontrakcijām, izraisa to ātrāku nogurumu.
Palielinoties nerva vai muskuļa stimulācijas biežumam, palielinās gludas stingumkrampju amplitūda. Maksimālais stingumkrampju skaits ir nosaukts optimāls. Stingumkrampju palielināšanās skaidrojama ar Ca2+ uzkrāšanos hialoplazmā. Turpinot palielināt nervu stimulācijas biežumu (apmēram 100 Hz), muskuļi atslābina, jo neiromuskulārajās sinapsēs veidojas bloks ierosmes vadīšanā - Vvedenska pesīms(kairinājuma biežums pesimistisks) (skat. 26. att.). Vvedensky pessimum var iegūt arī ar tiešu, bet biežāku muskuļa stimulāciju (apmēram 200 impulsi/s), tomēr eksperimenta tīrības labad ir jābloķē neiromuskulārās sinapses. Ja pēc pesimas rašanās stimulācijas biežums tiek samazināts līdz optimālajam, tad muskuļu kontrakcijas amplitūda acumirklī palielinās - pierādījums tam, ka pessims nav muskuļu noguruma vai enerģijas resursu izsīkuma rezultāts.
Dabiskos apstākļos atsevišķas muskuļu šķiedras nereti saraujas zobainā stingumkrampju režīmā, tomēr visa muskuļa kontrakcija atgādina gludu stingumkrampju, jo to kontrakcija ir asinhrona.
FEDERĀLĀ IZGLĪTĪBAS AĢENTŪRA
PENZAS VALSTS PEDAGOĢISKĀ UNIVERSITĀTE, NOSAUKTA V.G. BELINSKIS
Pieņemts Dabas ģeogrāfijas fakultātes Akadēmiskās padomes sēdē Protokols Nr.___ datēts ar "___" _________ 2006.g.
Fakultātes dekāns ________________
L.V. Krivošejeva ES APSTIPRINU
Akadēmisko lietu prorektors
______________________________
M.A. Pjatins
IZGLĪTĪBAS DARBA PROGRAMMA
disciplīnā "Muskuļu kontrakcijas bioķīmija"
par specialitāti
020208 (012300) - "Bioķīmija"
Dabas ģeogrāfijas fakultāte
Bioķīmijas katedra
Penza, 2006
DISCIPLINAS VALSTS IZGLĪTĪBAS STANDARTA PRASĪBAS
Rādītājs
KVALIFIKĀCIJAS PRASĪBAS
Bioķīmiķa speciālista sagatavošana notiek bioloģiskajās fakultātēs vai katedrās, bioķīmijas katedrās. Bioķīmiķa speciālista galvenās izglītības programmas īstenošana jānodrošina skolotājiem ar pamatizglītību un/vai darba pieredzi un publikācijām mācījamo disciplīnu profilā, sistemātiski veicot zinātnisko un zinātniski metodisko darbu, kas apstiprināts ar publikācijām. Skolotāju īpatsvaram ar akadēmiskajiem grādiem un nosaukumiem jābūt vismaz 67%. Speciālo disciplīnu skolotājiem, kā likums, ir jābūt akadēmiskajam grādam un pieredzei attiecīgajā profesionālajā jomā.
DISCIPLĪNAS MĒRĶI UN MĒRĶI
Kursā "Muskuļu kontrakciju bioķīmija" jāiepazīstina studenti ar muskuļu molekulārās struktūras un funkcionēšanas pamatiem normālos un patoloģiskos apstākļos, kā arī bioloģisko procesu īpatnībām, kas tajos notiek fizisko vingrinājumu un sporta laikā.
Muskuļu kontrakcijas bioķīmija ir bioķīmijas nozare un atrodas teorētiskās un eksperimentālās fizikas, molekulārās bioloģijas, cilvēka fizioloģijas un medicīnas disciplīnu krustpunktā.
Kursa saturs sastāv no tēmām par olbaltumvielu uzbūves un darbības mehānisma īpatnībām muskuļu audos, muskuļu aktivitātes energoapgādi un bioķīmisko procesu norisi normālos apstākļos, kā arī slodzes laikā un patoloģiju. Šī kursa apguve ļauj topošajiem speciālistiem izprast ķermeņa muskuļu audos notiekošo procesu būtību un nozīmi molekulārā līmenī.
Kursa mērķi: apvienot fundamentālo informāciju par cilvēka bioķīmiju un iespēju šīs zināšanas izmantot fiziskās audzināšanas praksē.
Kursa mērķi:
Sniegt mūsdienīgu informāciju par vielmaiņas īpatnībām muskuļu aktivitātes laikā;
Atklājiet bioķīmiskos pamatus:
izmaiņas organismā fiziskās slodzes laikā
noguruma procesi
atveseļošanās procesi
apmācības pielāgojumi
apmācības metodes
apmācības procesa efektivitāte
sporta sniegums
motorisko īpašību un sportistu izturības attīstība
ēdiens
uzraudzīt sportistu stāvokli
Programma ir sastādīta saskaņā ar Valsts augstākās profesionālās izglītības standartu studentiem, kas uzņemti specialitātē 020208 (012300) "Bioķīmija".
Atbilstoši šīs specialitātes mācību programmai kursam "Muskuļu kontrakcijas bioķīmija" ir atvēlētas 68 stundas, no kurām 34 stundas auditorijai un 34 stundas patstāvīgajam darbam. No 34 stundām auditorijā 34 stundas ir lekcijas. Kurss nodrošina kredītpunktu.
MĀCĪBU LAIKA SADALĪJUMS PA SEMESTRIEM UN MĀCĪBU VEIDIEM
semestris
DISCIPLINAS SATURS
Ievads
Muskuļu kontrakcijas bioķīmijas priekšmets. Kursa mērķi un saturs. Īss vēsturisks pārskats. Muskuļu kontrakcijas bioķīmijas vērtība kā bioķīmiķu apmācības priekšmets. Muskuļu kontrakcijas bioķīmijas pozīcija dabaszinātņu vispārējā sistēmā.
Citoplazmas muskuļu un skeleta sistēma
Citoskeleta kontraktilie proteīni.
Šķiedru struktūru veidi: mikrofilamenti, mikrotubulas, starppavedieni. To struktūra un funkciju regulējums.
Skropstu struktūra un kustība. Baktēriju motora aparāts.
Kolagēns
kolagēna veidi. Kolagēna molekulu struktūras iezīmes. Kolagēna sintēze. Iedzimtas slimības, ko izraisa kolagēna anomālijas.
Muskuļu audu struktūra un ķīmiskais sastāvs
Muskuļu audu klasifikācija.
Svītrotā muskuļa morfoloģiskā organizācija. Muskuļu šķiedras struktūra. Svītrotā muskuļa ķīmiskais sastāvs. Muskuļu proteīni, kas veido sarkoplazmu: mioglobīns, parvalbumīns. Kontrakcijas olbaltumvielas: miozīns, aktīns, aktomiozīns, tropomiozīns, troponīns T, troponīns es, troponīns C, - un -aktīns. Slāpekļa ekstraktvielas, kas nesatur olbaltumvielas. slāpekli nesaturošas vielas.
Sirds muskuļa un gludo muskuļu ķīmiskā sastāva iezīmes.
Muskuļu audu ķīmiskā sastāva izmaiņas ontoģenēzē.
5. Funkcionālā muskuļu bioķīmija
Muskuļu kontrakcijas bioķīmiskais cikls. Muskuļu kontrakcijas un relaksācijas regulēšana: šķērssvītroto muskuļu aktīna regulēšana, gludo muskuļu miozīna regulēšana. Kalcija jonu un modulējošo proteīnu loma.
6. Bioenerģētiskie procesi muskuļu darbības laikā
ATP resintēzes anaerobie (miokināzes, kreatīnfosfokināzes un glikolītiskā) un aerobā (oksidatīvā fosforilēšanās) ceļi muskuļu aktivitātes laikā. Dažādu ATP resintēzes ceļu attiecība dažādas jaudas un intensitātes muskuļu aktivitātes laikā. Dažādu enerģijas avotu ieslēgšanas secība.
7. Vingrošanas un sporta bioķīmija
Skābekļa transporta bioķīmiskās īpašības un tā nogulsnēšanās muskuļos. Skābekļa patēriņš darbā, skābekļa deficīts un skābekļa parāds. Vingrošanas enerģijas izmaksas un skābekļa patēriņš.
Bioķīmiskās izmaiņas, kas treniņu ietekmē notiek muskuļu sistēmā, iekšējos orgānos, asinīs un centrālajā nervu sistēmā. Bioķīmiskās izmaiņas standarta un maksimālā darba laikā, ar nogurumu, atpūtas periodā. Bioķīmisko izmaiņu secība treniņa un detrenēšanas laikā. Bioķīmiskās izmaiņas pārtrenēšanās laikā.
Sporta vingrinājumu klasifikācijas bioķīmiskais pamatojums pēc relatīvās jaudas. Bioķīmiskās izmaiņas organismā ciklisko (skriešana, peldēšana, riteņbraukšana, slēpošana un slidošana) un aciklisko (svarcelšana, bokss, vingrošana, cīņa) laikā.
8. Bioķīmiskās izmaiņas muskuļos patoloģijā
Aptuvenais stundu sadalījums pa tēmām
p/n
KONTROLES DARBU PIEMĒRI
1. Muskuļu audu veidi. Muskuļu struktūra. Muskuļu kontrakcijas bioķīmija. Metabolisms muskuļu aktivitātes laikā - Tests.
Literatūra
Galvenais:
Papildus:
2. Bioķīmiskās izmaiņas organismā, veicot dažāda jaudas un ilguma vingrinājumus. Noguruma un atveseļošanās bioķīmiskie faktori. Bioķīmiskās adaptācijas modeļi sporta treniņu laikā. Izturības bioķīmiskie pamati, ātruma-spēka īpašības, sportiskais sniegums. - Problēmu risināšana.
Literatūra
Galvenais:
1. Alberts B., Bray D., Lewis J., Ruff M., Roberts K., Watson J. Molecular biology of the cell. 3 sējumos. 2. izd. - M.: Mir, 1994
2. Berezovs T.T., Korovkins B.F. Bioloģiskā ķīmija. - M.: Medicīna, 2002
3. Marie R., Grenner D., Meyes P., Rodwell V. Cilvēka bioķīmija: Per. no angļu valodas. - M.: Mir, 1993
4. Bioķīmija / Red. Menšikova V.V., Volkova N.I. - M .: Fiziskā kultūra un sports, 1986
5. Volkovs N.I. Muskuļu aktivitātes bioķīmija. - M .: Olimpiskais sporta veids, 2001.
6. Jakovļevs N.N. Sporta bioķīmija. - M .: Fiziskā kultūra un sports, 1974
7. Jakovļevs N.N. Kustības ķīmija. - L .: Nauka, 1983
Papildus:
1. Cilvēka fizioloģija / Kositsky G.I. – M.: Medicīna, 1985, 544lpp.
2. Bioķīmija un molekulārā bioloģija / Elliots V., Eliots D.; Per. no angļu valodas: O.V. Dobrynina un citi; Ed. A.I. Arčakova - M .: MAIS "Nauka / Interperiodika", 2002, 446s.
3. Lēningers A. Bioķīmija. T. 1 - 3. M .: Mir, 1985
ZINĀŠU GALĪGĀS KONTROLES FORMA - TESTS
OFFSET JAUTĀJUMU SARAKSTS PIEMĒRS
Metabolisma iezīmes muskuļu darbības laikā.
Muskuļu un muskuļu šķiedru veidi. Muskuļu šķiedru strukturālā organizācija. Muskuļu audu ķīmiskais sastāvs.
Strukturālās un bioķīmiskās izmaiņas muskuļos kontrakcijas un relaksācijas laikā. Muskuļu kontrakcijas molekulārais mehānisms.
Muskuļu kontrakcijas un relaksācijas regulēšana: šķērssvītroto muskuļu aktīna regulēšana, gludo muskuļu miozīna regulēšana. Kalcija jonu un modulējošo proteīnu loma.
Enerģijas ražošanas mehānismu vispārīgie raksturojumi. Kreatīnfosfokināze, glikolītiskā, miokināze, ATP resintēzes aerobie mehānismi. Dažādu ATP resintēzes ceļu attiecība dažādas jaudas un intensitātes muskuļu aktivitātes laikā. Dažādu enerģijas avotu ieslēgšanas secība.
Bioķīmisko procesu izmaiņu vispārējais virziens muskuļu darbības laikā. Skābekļa transportēšana uz strādājošiem muskuļiem un tā patēriņš muskuļu darbības laikā.
Bioķīmiskās izmaiņas atsevišķos orgānos un audos muskuļu darba laikā. Fizisko vingrinājumu klasifikācija pēc bioķīmisko izmaiņu rakstura muskuļu darba laikā.
Noguruma bioķīmiskie faktori.
Atveseļošanās procesu bioķīmiskie pamati.
Faktori, kas ierobežo cilvēka fizisko veiktspēju. Sportista aerobās un anaerobās veiktspējas rādītāji.
Treniņu ietekme uz sportistu sniegumu. Vecums un sportiskais sniegums.
Ātruma-stiprības īpašību bioķīmiskās īpašības. Sportistu ātruma-spēka treniņu metožu bioķīmiskie pamati.
Izturības bioķīmiskie faktori. Treniņu metodes, kas veicina izturības attīstību.
Fiziskās aktivitātes, adaptācijas un treniņu efekts. bioķīmiskās adaptācijas attīstības modeļi un apmācības principi. Adaptīvo izmaiņu specifika organismā treniņa laikā.
Adaptīvo izmaiņu atgriezeniskums treniņa laikā. Adaptīvo izmaiņu secība treniņa laikā.
Apmācības efektu mijiedarbība treniņa laikā. Adaptācijas cikliskā attīstība apmācības procesā.
Sportistu racionāla uztura principi. Organisma enerģijas patēriņš un tā atkarība no veiktā darba. Uzturvielu līdzsvars sportistu uzturā.
Atsevišķu pārtikas ķīmisko komponentu loma muskuļu aktivitātes nodrošināšanā. Uztura bagātinātāji un ķermeņa svara regulēšana.
Bioķīmiskās kontroles uzdevumi, veidi un organizācija. Pētījuma objekti un bioķīmiskie pamatparametri. Galvenie asins un urīna sastāva bioķīmiskie rādītāji, to izmaiņas muskuļu darbības laikā. Organisma energoapgādes sistēmu attīstības bioķīmiskā kontrole muskuļu aktivitātes laikā.
Bioķīmiskā kontrole pār treniņu līmeni, sportista ķermeņa nogurumu un atjaunošanos. Dopinga kontrole sportā.
II. Referātu sagatavošana: Dažādu sporta veidu bioķīmiskie pamati.
Plāns:
energosistēmu savienošana un pielāgošana treniņu laikā
aerobo un anaerobo procesu jauda un jauda
bioķīmiskās izmaiņas organismā
noguruma un atveseļošanās procesi
adaptīvo izmaiņu specifika treniņa laikā
pārtikas bioķīmija
DISCIPLINAS IZGLĪTĪBAS UN METODOLOĢISKAIS ATBALSTS
Alberts B., Bray D., Lewis J., Ruff M., Roberts K., Watson J. Molecular biology of the cell. 3 sējumos. 2. izd. - M.: Mir, 1994
Berezovs T.T., Korovkins B.F. Bioloģiskā ķīmija. - M.: Medicīna, 2002
Lehningers A. Bioķīmija. T. 1 - 3. M .: Mir, 1985
Marie R., Grenner D., Meyes P., Rodwell W. Cilvēka bioķīmija: Per. no angļu valodas. - M.: Mir, 1993
Bioķīmija un molekulārā bioloģija / Elliots V., Eliots D.; Per. no angļu valodas: O.V. Dobrynina un citi; Ed. A.I. Arčakova - M .: MAIS "Nauka / Interperiodika", 2002, 446s.
Bioķīmija / Red. Menšikova V.V., Volkova N.I. - M .: Fiziskā kultūra un sports, 1986
PAPILDU LITERATŪRA
Volkovs N.I. Muskuļu aktivitātes bioķīmija. - M .: Olimpiskais sporta veids, 2001.
Cilvēka fizioloģija / Kositsky G.I. – M.: Medicīna, 1985, 544lpp.
Jakovļevs N.N. Sporta bioķīmija. - M .: Fiziskā kultūra un sports, 1974
Jakovļevs N.N. Kustības ķīmija. - L .: Nauka, 1983
Bioķīmijas katedras sēdē tika apspriesta un apstiprināta mācību programma disciplīnai "Muskuļu kontrakcijas bioķīmija" specialitātei 0202028 (012300) - "Bioķīmija".
Protokols Nr._____ datēts ar "____" _____________ 2006.g
Galva Bioķīmijas katedra
Bioloģijas zinātņu doktors, profesors __________________________________________ М.Т. Gengin
(paraksts)
Apstiprinājusi Dabas ģeogrāfijas fakultātes Metodiskā padome
Protokols Nr.____________ datēts ar "_____" _______________ 2006.g
Metodiskās padomes priekšsēdētājs
Dabas ģeogrāfijas fakultāte,
Tehnisko zinātņu kandidāts, asociētais profesors ______________________ O.V. Zorkins
(paraksts)
Sastādīja:
Cand. biol. Zinātnes, asociētā profesore Petrušova O.P. _________________________________
(paraksts)
3. NODAĻA. FUNKCIONĀLĀ MUSKUĻU BIOĶĪMIJA
3.1. Muskuļu kontrakcijas mehānisms
Neskatoties uz muskuļu sistēmas daudzfunkcionalitāti, muskuļu galvenā funkcija ir motora darbības, tas ir, kontrakcijas un relaksācijas, īstenošana. Muskuļu kontrakcija ir sarežģīts mehāniski ķīmisks process, kura laikā ATP hidrolītiskā sadalīšanās ķīmiskā enerģija tiek pārvērsta mehāniskajā enerģijā. Apskatīsim mugurkaulnieku šķērssvītroto muskuļu kontrakcijas procesa strukturālo pamatu, jo šis process ir izpētīts vispilnīgāk. Kā minēts, šķērssvītrotā muskuļa kontraktilā sistēma sastāv no pārklājošiem proteīna pavedieniem, kas slīd viens pret otru (sk. 9. att., A).
Saskaņā ar E. Hakslija un R. Nīdergerkes, kā arī X. Hakslija un J. Hensona piedāvāto modeli, miofibrilām saraujoties, viena pavedienu sistēma iekļūst citā, tas ir, pavedieni sāk slīdēt viens pāri, kā tā bija, kas ir muskuļu kontrakcijas cēlonis.
Kontrakcijas notiek ATP hidrolīzes laikā atbrīvotās enerģijas dēļ. Svītrotajā muskulī kontrakcijas ir atkarīgas no Ca 2+ jonu koncentrācijas, ko savukārt regulē sarkoplazmatiskais tīklojums – specializēta membrānu sistēma, kas miera stāvoklī uzkrāj Ca 2+ un atbrīvo to, kad uz muskuļu tiek pievadīts nervu impulss. muskuļu šķiedra (sk. 11. att., A, B).
1) miozīna "galva" var hidrolizēt ATP par ADP un H 3 PO 4 (P i), bet nenodrošina hidrolīzes produktu izdalīšanos. Tāpēc šim procesam ir drīzāk stehiometrisks, nevis katalītisks raksturs (sk. 10. att., a);
3) šī mijiedarbība nodrošina ADP un H 3 RO 4 izdalīšanos no aktīna-miozīna kompleksa. Akomiozīna saitei ir viszemākā enerģija 45° leņķī, tāpēc miozīna leņķis ar fibrila asi mainās no 90° līdz 45° (aptuveni) un aktīns virzās (par 10-15 nm) sarkomēra centra virzienā (sk. 10. att., c ) (“gājiens” pēc airu laivas modeļa 9. att. B);
Rīsi. 9. Mugurkaulnieku skeleta muskuļu organizācija
un muskuļu kontrakcijas mehānisms
4) jauna ATP molekula saistās ar miozīna-F-aktīna kompleksu (skat. 10. att., d);
5) miozīna-ATP kompleksam ir zema afinitāte pret aktīnu, un tāpēc notiek miozīna (ATP) “galvas” atdalīšanās no F-aktīna. Pēdējais posms faktiski ir relaksācija, kas nepārprotami ir atkarīga no ATP saistīšanās ar aktīna-miozīna kompleksu (sk. 10.e att.). Pēc tam cikls atsākas.
Rīsi. desmit. Muskuļu kontrakcijas bioķīmiskais cikls
Cikls atkārtojas tik ilgi, kamēr ir ATP. Katrs 500 biezu pavedienu miozīna "galviņu" "gājiens" izraisa 10 nm nobīdi. Spēcīgu kontrakciju laikā insulta ātrums ir aptuveni 5 reizes sekundē. Ar katru ATP hidrolīzes ciklu miozīna “galvas” mijiedarbojas ar jaunām aktīna molekulām, kā rezultātā notiek miozīna un aktīna pavedienu savstarpēja slīdēšana, tas ir, muskuļu šķiedru kontrakcija.
3.2. Muskuļu kontrakcijas un relaksācijas regulēšana
Jebkura muskuļa kontrakcija notiek saskaņā ar iepriekš aprakstīto vispārējo mehānismu. Dažādu orgānu muskuļu šķiedrām var būt dažādi kontrakcijas un relaksācijas regulēšanas molekulārie mehānismi, taču Ca 2+ joniem vienmēr ir galvenā regulējošā loma. Konstatēts, ka miofibrilām piemīt spēja mijiedarboties ar ATP un tā klātbūtnē sarauties tikai tad, ja vidē ir noteikta kalcija jonu koncentrācija. Vislielākā saraušanās aktivitāte tiek novērota Ca 2+ jonu koncentrācijā aptuveni 10–6–10–5 M. Koncentrācijai nokrītot līdz 10–7 M vai mazāk, muskuļu šķiedras zaudē spēju saīsināties un attīstīt spriedzi klātbūtnē. no ATP.
Saskaņā ar mūsdienu koncepcijām miera stāvoklī esošajos muskuļos (miofibrilās un starpfibrilārajā telpā) Ca 2+ jonu koncentrācija tiek uzturēta zem sliekšņa vērtības, to saistīšanās rezultātā ar sarkoplazmatiskā retikuluma struktūrām (kanāliņiem un pūslīšiem) un tā tālāk. -ko sauc par T-sistēmu, kurā piedalās īpašs Ca 2+ -saistošais proteīns, ko sauc par kalsequestrīnu, kas ir daļa no šīm struktūrām.
Ca 2+ jonu saistīšanās ar plašu sarkoplazmatiskā retikuluma kanāliņu un cisternu tīklu nav vienkārša adsorbcija. Tas ir aktīvs fizioloģisks process, kas tiek veikts, pateicoties enerģijai, kas izdalās sarkoplazmas retikuluma ATP Ca 2+ atkarīgās ATPāzes sadalīšanās laikā. Šajā gadījumā tiek novērota ļoti savdabīga aina: Ca 2+ jonu izvadīšanas ātrumu no starpfibrilārās telpas stimulē tie paši joni. Kopumā šādu mehānismu sauca par "kalcija sūkni" pēc analoģijas ar fizioloģijā labi zināmo nātrija sūkni (sk. 11. att., B).
Iespēja, ka dzīvs muskulis atrodas atslābinātā stāvoklī, ja tajā ir pietiekami augsta ATP koncentrācija, ir izskaidrojama ar Ca 2+ jonu koncentrācijas samazināšanos miofibrilu apkārtējā vidē kalcija iedarbības rezultātā. sūknis, zem robežas, pie kuras joprojām ir iespējama ATPāzes aktivitātes izpausme un šķiedras aktomiozīna struktūru kontraktilitāte. Muskuļu šķiedras strauja kontrakcija, kad to stimulē nervs (vai elektriskā strāva), ir pēkšņas membrānu caurlaidības maiņas rezultāts, kā rezultātā izdalās noteikts Ca 2+ daudzums. joni no sarkoplazmatiskā tīkla un T-sistēmas tvertnēm un kanāliņiem nonāk sarkoplazmā (sk. 11. att., A, B).
Kā minēts, aktomiozīna sistēmas “jutība” pret Ca 2+ joniem (tas ir, aktomiozīna spējas šķelties ATP un saraušanās ATP klātbūtnē zudums, Ca 2+ jonu koncentrācijai samazinoties līdz 10 -7 M) ir saistīts ar troponīna proteīna klātbūtni kontraktilajā sistēmā (uz F-aktīna pavedieniem), kas saistīts ar tropomiozīnu. Troponīna-tropomiozīna kompleksā Ca 2+ joni precīzi saistās ar troponīnu (troponīna C apakšvienība pēc īpašībām ir līdzīga kalmodulu-

Rīsi. vienpadsmit. Muskuļu kontrakciju regulēšana
labi). Ca 2+ jonu saistīšanās izraisa konformācijas izmaiņas troponīna molekulā, kas acīmredzot noved pie visa troponīna-tropomiozīna stieņa nobīdes un aktīna aktīvo vietu atbloķēšanas, kas var mijiedarboties ar miozīnu, veidojot saraušanās kompleksu un aktīvu. Mg2+-ATPāze. Tas sāk muskuļu kontrakcijas ciklu (sk. 11.B attēlu).
Aktīna pavedienu veicināšanā gar miozīna pavedieniem, pēc E. Hakslija domām, svarīga loma ir šķērsvirziena tiltiem, kas īslaicīgi noslēdzas starp pavedieniem, kas ir miozīna molekulu "galvas". Tātad, jo lielāks ir aktīna pavedieniem piesaistīto tiltu skaits noteiktā brīdī, jo lielāks ir muskuļu kontrakcijas spēks.
Visbeidzot, ja ierosme apstājas, Ca 2+ jonu saturs sarkoplazmā samazinās (kalcija sūknis), kā rezultātā Ca 2+ komplekss ar troponīnu C disociējas, troponīns atjauno sākotnējo konformāciju, tiek bloķēta miozīna saistīšanās vieta uz aktīna. , tas ir, miozīna pavedienu “galvas” pārstāj pievienoties aktīna pavedieniem. ATP klātbūtnē muskuļi atslābinās un tā garums sasniedz sākotnējo garumu. Ja ATP padeve apstājas (anoksija, inhalācijas saindēšanās vai nāve), muskuļi nonāk stingrības stāvoklī. Gandrīz visi biezo (miozīna) pavedienu šķērseniskie tilti ir pievienoti plāniem aktīna pavedieniem, kā rezultātā muskulis ir pilnībā nekustīgs.
4. NODAĻA. MUSKUĻU AKTIVITĀTES BIOENERĢIJA
4.1. Sistēmu un mehānismu vispārīgie raksturojumi
muskuļu aktivitātes energoapgāde
Kā parādīts 3. nodaļā, ATP ir tūlītējs enerģijas avots muskuļu darbībai. Enerģijas izdalīšanās notiek ATP molekulas fermentatīvās hidrolīzes laikā par ADP un ortofosfātu:
Ca 2+ -ATPāze
ATP + H 2 O ADP + H 3 RO 4.
ΔQ = 7,3 kcal jeb 30 kJ
Ķīmiskā enerģija muskuļu kontrakcijas procesā tiek pārvērsta muskuļu mehāniskajā darbā, un relaksācijas laikā nodrošina aktīvu Ca 2+ transportēšanu uz sarkoplazmas tīklu. Skeleta muskuļos tiek patērēts liels ATP daudzums Na + -K + -ATPāzes darbam, kas uztur noteiktu Na + un K + jonu koncentrāciju muskuļos, radot elektroķīmisko potenciālu uz sarkolemmas.
Tādējādi, lai nodrošinātu muskuļu šūnu ar tās saraušanās aparātu ar pietiekamu enerģijas daudzumu ATP formā, ir nepieciešama nepārtraukta šī savienojuma resintēze.
ATP saturs muskuļos ir nenozīmīgs un ir aptuveni 5 mmol ∙ kg -1 no neapstrādāta audu masas (0,25–0,40%). Tas tiek uzturēts relatīvi nemainīgā līmenī, jo ATP koncentrācijas palielināšanās muskuļos izraisa miozīna ATPāzes inhibīciju, kas novērš saķeres veidošanos starp aktīna un miozīna pavedieniem miofibrilās un muskuļu kontrakciju, un tās samazināšanos zem 2 mmol ∙ kg -1 neapstrādātas audu masas noved pie Ca 2+ sūkņa darbības un muskuļu relaksācijas procesa pārkāpuma tīklā. ATP rezerves muskuļu šķiedrās spēj nodrošināt intensīva darba veikšanu tikai ļoti īsu laiku - 0,5–1,5 s, jeb 3–4 vienreizējas maksimālā spēka kontrakcijas. Turpmākais muskuļu darbs tiek veikts, pateicoties ātrai ATP atgūšanai (resintēzei) no tā sabrukšanas produktiem un enerģijas daudzumam, kas izdalījās sabrukšanas laikā:
ADP + H 3 RO 4 + ΔQ → ATP.
Fosfāta pievienošanas reakciju sauc par fosforilēšanu, un reakciju, kad to pārnes no vienas vielas uz citu, sauc par refosforilēšanu.


Rīsi. 12. Enerģijas vielmaiņa muskuļu audos
Enerģijas avoti ATP resintēzei skeleta muskuļos un citos audos ir ar enerģiju bagātas fosfātus saturošas vielas, kas atrodas audos (kreatīna fosfāts, ADP) vai veidojas glikogēna, taukskābju un citu enerģijas substrātu (piemēram, metabolītu) katabolisma laikā. difosfoglicerīnskābes un fosfopirovīnskābes), kā arī protonu (H +) gradienta enerģija pāri mitohondriju membrānai, kas rodas dažādu vielu aerobās oksidācijas rezultātā.
Atkarībā no tā, kurš bioķīmiskais process nodrošina enerģiju ATP molekulu veidošanai, ir četri mehānismi jeb ceļi ATP resintēzei audos (sk. 12. att.). Katram mehānismam ir savas vielmaiņas un bioenerģētiskās īpašības. Muskuļu darba enerģijas apgādē tiek izmantoti dažādi mehānismi atkarībā no veiktā vingrinājuma intensitātes un ilguma.
ATP resintēzi var veikt reakcijās, kas notiek bez skābekļa līdzdalības (anaerobie mehānismi) vai ar inhalējamā skābekļa līdzdalību (aerobais mehānisms).
Normālos apstākļos ATP resintēze audos notiek pārsvarā aerobā veidā, un intensīvas muskuļu darbības laikā, kad ir apgrūtināta skābekļa piegāde muskuļiem, audos palielinās arī ATP resintēzes anaerobie mehānismi. Cilvēka skeleta muskuļos ir identificēti trīs ATP resintēzes veidi anaerobā un viens aerobais ceļš (sk. 13. att.).
Anaerobie mehānismi ietver:
1) kreatīnfosfokināzes (fosfogēns vai alaktāts) mehānisms, kas nodrošina ATP resintēzi, pateicoties pārfosforilēšanai starp kreatīna fosfātu un ADP;
2) glikolītiskais (laktāta) mehānisms, kas nodrošina ATP resintēzi muskuļu glikogēna vai glikozes līmeņa asinīs fermentatīvā anaerobā sadalīšanās procesā, kas beidzas ar pienskābes veidošanos, un tāpēc tiek saukts par laktātu;
3) miokināzes mehānisms, kas resintezē ATP, pateicoties refosforilācijas reakcijai starp divām ADP molekulām, piedaloties enzīmam miokināzei (adenilātkināzei).

Rīsi. 13. ATP resintēzes mehānismi muskuļos
(ietvars parāda enerģijas substrātus
un iezīmēja mehānismu nosaukumus)
ATP resintēzes aerobais mehānisms galvenokārt ietver oksidatīvās fosforilēšanas reakcijas, kas notiek mitohondrijās. Aerobās oksidācijas enerģētiskie substrāti ir glikoze, taukskābes, daļēji aminoskābes, kā arī glikolīzes starpprodukti - pienskābe, taukskābju oksidācijas - ketonu ķermeņi.
Katram mehānismam ir atšķirīgas enerģijas iespējas, kuras raksturo šādi enerģijas ražošanas mehānismu novērtēšanas kritēriji: maksimālā jauda, izvēršanas ātrums, vielmaiņas kapacitāte un efektivitāte. Maksimālā jauda ir augstākais ATP ražošanas ātrums noteiktā vielmaiņas procesā. Tas ierobežo šī mehānisma veiktā darba maksimālo intensitāti. Izvēršanas ātrumu aprēķina pēc laika, kad no darba sākuma tiek sasniegta noteiktā ATP atkārtotas sintēzes ceļa maksimālā jauda. Metabolisma kapacitāte atspoguļo kopējo ATP daudzumu, ko var iegūt noteiktā resintēzes mehānismā enerģijas substrātu rezervju lieluma dēļ; jauda ierobežo veicamā darba apjomu. Metabolisma efektivitāte ir tā enerģijas daļa, kas uzkrājas ATP makroerģiskajās saitēs; tas nosaka veiktā darba efektivitāti un tiek novērtēts pēc kopējās veiktspējas koeficienta (COP) vērtības, kas ir visas iztērētās lietderīgās enerģijas attiecība pret tās kopējo šajā vielmaiņas procesā izdalīto daudzumu.
Kopējā efektivitāte vielmaiņas procesu enerģijas pārveidošanā mehāniskajā darbā (E m) ir atkarīga no diviem rādītājiem: a) vielmaiņas transformāciju laikā atbrīvotās enerģijas pārvēršanas efektivitāte atkārtoti sintezēto makroenerģētisko fosfora savienojumu (ATP) enerģijā, tas ir, fosforilēšanas efektivitāte (E f); b) ATP pārveidošanas efektivitāte mehāniskajā darbā, tas ir, ķīmiskās mehāniskās savienojuma efektivitāte (E e):
E m = (E f / E e) × 100.
Ķīmijmehāniskās konjugācijas efektivitāte procentos no aerobās un anaerobās vielmaiņas ir aptuveni vienāda un sastāda 50%, savukārt fosforilācijas efektivitāte ir visaugstākā alaktiskajā anaerobajā procesā - aptuveni 80% un zemākā - anaerobajā glikolīzē - vidēji 44%, aerobā procesā tas ir aptuveni 60%.
Kreatīnfosfokināzes, glikolītisko un aerobo muskuļu aktivitātes energoapgādes mehānismu salīdzinošie raksturojumi atbilstoši vērtēšanas kritērijiem parādīti 3. tabulā.
3. tabulā parādīts, ka kreatīnfosfokināzes un glikolītiskajiem mehānismiem ir liela ATP veidošanās maksimālā jauda un efektivitāte, bet īss maksimālās jaudas saglabāšanas laiks un maza jauda, pateicoties mazajām enerģijas substrātu rezervēm. Aerobajam mehānismam ir gandrīz trīs reizes mazāka maksimālā jauda salīdzinājumā ar kreatīnfosfokināzi, taču tā tiek uzturēta ilgu laiku, kā arī gandrīz neizsmeļama jauda, pateicoties lielajām enerģijas substrātu rezervēm ogļhidrātu, tauku un daļēji olbaltumvielu veidā. Tātad, pateicoties tauku rezervēm, ķermenis var nepārtraukti strādāt 7-10 dienas, savukārt anaerobo enerģijas ražošanas mehānismu enerģijas substrātu rezerves ir mazāk nozīmīgas.
Apmācības dokuments
... UDC (470)(082) BBC ... publicēšana labdarība M. P. Beļajevs. Atsevišķas ballītes publicēšana ... iespiests ... padoms kurā tas tika pieņemts risinājums turpina savu darbību trimdā. Neļaujoties ilūzijām ieslēgts... kvalitāte izglītojošs ieguvumi. Viņa...