ამინომჟავები ცილების სტრუქტურული კომპონენტებია. ცილის მოლეკულების სტრუქტურული ორგანიზაციის დონეები. პოლარული დადებითად დამუხტული
1.1. ცილები. ამინომჟავები -- ცილების სტრუქტურული კომპონენტები
1.2. ამინომჟავების სტრუქტურა და კლასიფიკაცია
1.3. ცილის მოლეკულების სტრუქტურული ორგანიზაციის დონეები
1.4. ცილების ფიზიკოქიმიური თვისებები
1.5. ცილების კლასიფიკაცია
1.6. ნახშირწყლები. ნახშირწყლების კლასიფიკაცია
1.7. ლიპიდები
1.8. ვიტამინები
1.8.1. ცხიმში ხსნადი ვიტამინები
1.8.2. წყალში ხსნადი ვიტამინები
1.8.3. ვიტამინის მსგავსი ნივთიერებები.
2.1. ფერმენტები და არაორგანული კატალიზატორები
2.2. ფერმენტების სტრუქტურა
2.3. კოენზიმები
2.4. ფერმენტების თვისებები
2.5. ფერმენტების ნომენკლატურა
2.6. ფერმენტების კლასიფიკაცია
2.7. ფერმენტების მოქმედების მექანიზმი
2.8. ფერმენტული აქტივობის ინჰიბირება
3.1. ნახშირწყლების ბიოლოგიური როლი
3.2. საჭმლის მომნელებელ ტრაქტში ნახშირწყლების გარდაქმნა
3.3. გლიკოგენის ბიოსინთეზი და დაშლა
3.4. გლუკოზის კატაბოლიზმის ძირითადი გზები
3.4.1. ანაერობული გლიკოლიზი
3.4.2. აერობული გლიკოლიზი (ჰექსოზა დიფოსფატის გზა)
3.4.3. ჰექსოზა მონოფოსფატის გზა
3.4.4. გლუკონეოგენეზი
4.1. ადამიანის სხეულის ძირითადი ლიპიდები და მათი ბიოლოგიური როლი.
4.2. ლიპიდების მონელება, ცხიმის რესინთეზი
4.3. სისხლის ლიპოპროტეინები
4.4. ოქსიდაცია უმაღლესი ცხიმოვანი მჟავები
4.5. გლიცეროლის დაჟანგვა
4.6. IVH ბიოსინთეზი ქსოვილებში
4.7. ქოლესტერინის გაცვლა
5.1. ცილების მონელება
5.2. ამინომჟავების გაფუჭება, დამპალი პროდუქტების განეიტრალება
5.3. ამინომჟავების მეტაბოლიზმი
5.4. ამიაკის განეიტრალების გზები
6.1. სასიგნალო მოლეკულები
6.2. ჰიპოთალამუსის ჰორმონები
6.3. ჰიპოფიზის ჰორმონები
6.4. ფარისებრი ჯირკვლის ჰორმონები
6.5. პარათირეოიდული ჯირკვლების ჰორმონები
6.6. გონადალური ჰორმონები
6.7. თირკმელზედა ჯირკვლის ჰორმონები
6.8. პანკრეასის ჰორმონები
1. შესავალი ბიოქიმიაში
2. ფერმენტები
3. ნახშირწყლების ცვლა
4. ლიპიდური ცვლა
5. ცილის ცვლა
6. ნივთიერებათა ცვლის რეგულირება
7. საგამოცდო კითხვები
1.1. ციყვები. ამინომჟავები ცილების სტრუქტურული კომპონენტებია
ცილები
პროტეინები არის აზოტის შემცველი, მაღალმოლეკულური ორგანული ნაერთები, რომლებიც შედგება ამინომჟავებისგან, რომლებიც დაკავშირებულია ჯაჭვებში პეპტიდური ბმების გამოყენებით და აქვთ რთული სტრუქტურული ორგანიზაცია.
იგივე ამინომჟავები გვხვდება სხვადასხვა სტრუქტურისა და ფუნქციის მქონე ცილებში. ცილის მოლეკულების ინდივიდუალობა განისაზღვრება ცილაში ამინომჟავების მონაცვლეობის თანმიმდევრობით.
ცილების დამახასიათებელი ნიშნები, რომლებიც განასხვავებენ მათ უჯრედის სხვა ორგანული ნაერთებისგან:
1. ცილები აზოტის შემცველი ნაერთებია, ისევე როგორც მრავალი სხვა უჯრედის კომპონენტი (ნუკლეინის მჟავები, ზოგიერთი ლიპიდი, ნახშირწყლები), მაგრამ სხვა ორგანული ნივთიერებებისგან განსხვავებით, აზოტის შემცველობა გაცილებით მაღალია - საშუალოდ 16 გრამი 100 გრამ ცილაზე.
2. ცილების სტრუქტურული ერთეულია L-სერიის ალფა ამინომჟავები.
3. ამინომჟავები დაკავშირებულია ცილებში პეპტიდური ბმების გამოყენებით, ქმნიან პოლიპეპტიდურ ჯაჭვს.
4.პროტეინებს აქვთ დიდი მოლეკულური წონა (20000-დან რამდენიმე მილიონ დალტონამდე).
5. ცილები განსხვავდებიან რთული სტრუქტურული ორგანიზაციით (მათ აქვთ პირველადი, მეორადი, მესამეული და მეოთხეული სტრუქტურა).
ცილების მრავალფეროვანი ფუნქციები:
1.კატალიტიკური. ბევრი ცილა არის ფერმენტი
2. მარეგულირებელი. ზოგიერთი ჰორმონი არის ცილა
3.სტრუქტურული. ცოცხალი უჯრედის ყველა სტრუქტურა შეიცავს ცილებს.
4. რეცეპტორი. ცილები რეცეპტორების აუცილებელი კომპონენტია და შეუძლიათ სხვა მოლეკულების ამოცნობა.
5. ტრანსპორტი. ისინი ატარებენ ცხიმებს, პიგმენტებს, ჰორმონებს, სამკურნალო ნივთიერებები, სხვადასხვა ჰიდროფობიური ნაერთები და ა.შ.
6. მხარდაჭერა. კოლაგენი, ძვლის ქსოვილის ცილები.
7.ენერგია. 1 გ ცილის დაჟანგვას თან ახლავს 17,6 კჯ ენერგიის გამოყოფა. ენერგიის 15% დღეში წარმოიქმნება ცილების დაშლის გამო.
8.კონტრაქტული. ამ ფუნქციას ასრულებენ კუნთოვანი ქსოვილის პროტეინები - აქტინი, მიოზინი.
9.ჰისტონის გენის მარეგულირებელი ფუნქცია.
10.იმუნოლოგიური. ანტისხეულები არის ცილები.
11. ჰემოსტატიკური. ფიბრინოგენი და სისხლის პლაზმის სხვა ცილები მონაწილეობენ სისხლის შედედების პროცესებში.
1.1.ამინომჟავები -- ცილების სტრუქტურული კომპონენტები
ამინომჟავები ჰეტეროფუნქციური ნაერთებია. ისინი წარმოადგენენ კარბოქსილის მჟავების წარმოებულებს, რომლებშიც წყალბადის ატომი ალფა მდგომარეობაში ჩანაცვლებულია ამინო ჯგუფით.
ცილების შემადგენელი ყველა ამინომჟავა ხასიათდება შემდეგი მახასიათებლები:
ა) ამინო ჯგუფის პოზიციის მიხედვით ისინი ა-ამინომჟავებია (ამინო ჯგუფი კარბოქსილის ჯგუფის გვერდით არის).
ბ) ისინი ყველა მიეკუთვნება L-სერიას, ვინაიდან ასიმეტრიული (ქირალური) ნახშირბადის ატომის ამინო ჯგუფი მარცხნივ იწერება. გამონაკლისი არის გლიცინი, რომელსაც არ აქვს ასიმეტრიული ნახშირბადის ატომი; მისი გვერდითი ჯაჭვი წარმოდგენილია წყალბადის ატომით.
გ) აქვთ ერთი და იგივე ზოგადი ფრაგმენტი და განსხვავდებიან მხოლოდ გვერდითი ჯაჭვის აგებულებით (R), რომლის თვისებები დიდწილად განსაზღვრავს თავად ამინომჟავების თვისებებს და იმ ცილებს, რომლებშიც ისინი შედის. სწორედ ეს განსხვავებები ფორმის, ზომისა და პოლარობის საშუალებას აძლევს ამინომჟავებს იყოს სამშენებლო ბლოკები, რომლებსაც ევოლუცია იყენებს ცილის სტრუქტურაზე მკაცრი მოთხოვნების დასაკმაყოფილებლად.
დ) ხსნარებში ნეიტრალური pH მნიშვნელობებით, ალფა ამინომჟავები ძირითადად გვხვდება შიდა მარილების სახით. ძლიერ მჟავე გარემოში ჭარბობს კათიონური ფორმა, ძლიერ ტუტე გარემოში – ანიონური ფორმა. (
IN სხვადასხვა ორგანიზმებიაღმოჩენილია მრავალი ამინომჟავა, რომელიც არ არის ცილების ნაწილი (ე.წ. არაცილოვანი ამინომჟავები). თუმცა, ყველა ცნობილი ორგანიზმი იყენებს ერთსა და იმავე 20 ამინომჟავას მათი ცილების შესაქმნელად; ფ. კრიკმა მათ "ჯადოსნური ოცი" უწოდა. მხოლოდ ისინი დაშიფრულია გენეტიკური კოდით.
მინერალები
მაკროელემენტების ჯგუფში შედის ისინი, რომელთა შემცველობა მშრალ ნივთიერებაში მერყეობს nּ10 -2-დან nּ10%-მდე. ეს არის C, O, H, N, S და P, რომლებიც ძირითადი ნივთიერებების მოლეკულური შემადგენლობის ნაწილია და Ca, Na, Cl, K, Mg, რომლებიც წარმოადგენს დამხმარე ქსოვილების, სისხლის, ლიმფის და სხვა ქსოვილების ნაწილს.
ულტრამიკროელემენტების ჯგუფში შედის ელემენტები, რომელთა შემცველობა nּ10 -5%-ზე დაბალია (Sb, Hg, Bi, Pb და ა.შ.).
დადგენილია, რომ ელემენტების უმეტესობა ბიოგენურია, დიდი მნიშვნელობა აქვს ბიოქიმიური სასიცოცხლო პროცესების ნორმალური განვითარების უზრუნველსაყოფად, ხოლო უმნიშვნელოვანესი ბიოგენური ელემენტები შედის IV (C); პერიოდული ცხრილის V (W, P) და VI (O, S) ჯგუფები. ელემენტების VII (Cl, J, Mn) და VIII (Fe, Co) ჯგუფები მონაწილეობენ მაღალი ბიოლოგიური ღირებულების მქონე ნივთიერებების წარმოქმნაში.
ნედლეულის მიკროელემენტის შემადგენლობა დამოკიდებულია ჰაბიტატზე ან ზრდაზე. დამოკიდებულია ცალკეული ელემენტების კონცენტრაციაზე გარემოდა საკვებში, მათ ხელმისაწვდომობაზე, ასევე შერჩევით უნარზე ცალკეული სახეობებიორგანიზმებს, იცვლება ასიმილაციის პროცესში ცალკეული ელემენტების გამოყენების ხარისხიც.
ციყვები
ორგანული ნივთიერებებიდან, რომლებიც ქმნიან ცოცხალ ორგანიზმებს, ბიოლოგიურად ყველაზე მნიშვნელოვანი და სტრუქტურით ყველაზე რთული ცილებია. სიცოცხლის თითქმის ყველა გამოვლინება (მონელება, გაღიზიანება, კონტრაქტურა, ზრდა და გამრავლება, მოძრაობა, მეტაბოლიზმი და ა.შ.) დაკავშირებულია ცილოვან ნივთიერებებთან. ცილები მნიშვნელოვან როლს ასრულებენ როგორც ცოცხალი ნივთიერების მშენებლობაში, ასევე მისი სასიცოცხლო პროცესების განხორციელებაში.
ცილოვანი ბუნების სპეციფიკური კატალიზატორები - ფერმენტები - აჩქარებენ ორგანიზმში მიმდინარე ქიმიურ რეაქციებს. სხვადასხვა ცილის ნაერთები ასრულებენ სატრანსპორტო ფუნქციას, ამარაგებენ ორგანიზმს ჟანგბადით და ნუტრიენტები. 1 გ ცილის საბოლოო პროდუქტად დაშლა ორგანიზმს 4,27 კკალ ენერგიას აძლევს.
ორგანოებიდან და ქსოვილებიდან გამოყოფილი პროტეინები გაცხელებისას წარმოქმნიან თეთრ ნალექს და აქვთ იგივე ფიზიკური თვისებები, რაც ცილებს. ქათმის კვერცხი. ამიტომაც დაიწყეს მათ ციყვის დარქმევა. სიტყვა "პროტეინის" სინონიმია სიტყვა "პროტეინი" (ბერძნულიდან "proteus" - პირველი, მთავარი).
ცილები არის სხვადასხვა ამინომჟავების მაღალი მოლეკულური წონის პოლიმერები. ნახ. 1 გვიჩვენებს სხვადასხვა ამინომჟავების ფორმულებს.
ნახ.1. ზოგიერთი ამინომჟავის ფორმულები.
ამინომჟავები იყოფა 2 ნაწილად დიდი ჯგუფები: გამოსაცვლელი და შეუცვლელი. ამინომჟავების უმეტესობა წარმოიქმნება ცხოველებისა და ადამიანების ორგანიზმში საკვების ცილების ჰიდროლიზისა და ბიოსინთეზის შედეგად. მაგრამ სულ მცირე რვა ამინომჟავა არ არის სინთეზირებული ორგანიზმში. ესენია ვალინი, ლეიცინი, იზოლეიცინი, თრეონინი, ლიზინი, ფენილალანინი, ტრიპტოფანი და მეთიონინი, რომლებსაც ესენციური ეწოდება. ცილებს, რომლებსაც აკლიათ ერთი ან მეტი ამინომჟავა, ბიოლოგიურად დეფიციტური ეწოდება. ცხოველური ცილები, მათ შორის წყლის ცილები, შეიცავს ყველა აუცილებელ ამინომჟავას.
ამინომჟავები, რომლებიც ქმნიან ცილას, ერთმანეთთან დაკავშირებულია პეპტიდური ბმებით, რომლებიც წარმოიქმნება ერთი ამინომჟავის ამინო ჯგუფსა და მეორის კარბოქსილის ჯგუფს შორის. ამ პროცესის მექანიზმი ნაჩვენებია ნახ. 2.
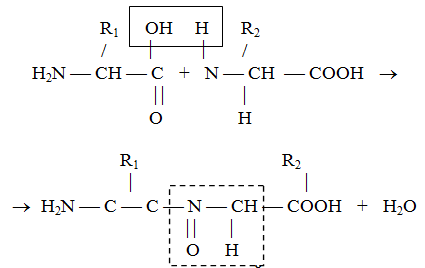
ბრინჯი. 2. ცილის პირველადი სტრუქტურის ფორმირება.
მიღებული პოლიპეპტიდები ყველა ცილის საფუძველია და მათში შემავალი ამინომჟავების სპეციფიკური თანმიმდევრობა ახასიათებს ცილის პირველად სტრუქტურას.
ამრიგად, რადგან ცილის მაკრომოლეკულები აგებულია მრავალი ასეული ამინომჟავისგან, ბუნებაში მათი იზომერების შეუზღუდავი რაოდენობაა და თითოეულ ცოცხალ არსებას შეიძლება ჰქონდეს თავისი უნიკალური ცილა.
პოლიპეპტიდურ ჯაჭვებს, თავის მხრივ, შეუძლიათ შეაერთონ მეორადი ცილის სტრუქტურების ჩამოყალიბება, ძირითადად ობლიგაციების გამო, რომლებიც წარმოიქმნება მათ შორის. სხვადასხვა ჯგუფებიპოლიპეპტიდები. ეს სქემატურად არის ნაჩვენები ნახ. 3.

ა) წყალბადის ბმების წარმოქმნა

ბ) პოლიპეპტიდური ჯაჭვიდან a-სპირალის წარმოქმნა
ბრინჯი. 3. ცილის მეორადი სტრუქტურის ფორმირების სქემა.
ცილის მოლეკულის პოლიპეპტიდური ჯაჭვების სივრცითი განლაგება განსაზღვრავს ცილის მოლეკულის მესამეულ სტრუქტურას.
თავად პროტეინები რთული სტრუქტურის მაღალმოლეკულური ნაერთებია, რომლებიც განსხვავდებიან როგორც ფიზიოლოგიური ფუნქციებით, ასევე ქიმიური თვისებები. საკვები ნედლეულის ცილები უპირატესად კოლოიდურ მდგომარეობაშია - გელისა და სოლის სახით, რაც განსაზღვრავს ცილოვანი ნივთიერებების თვისებების არასტაბილურობასა და ცვალებადობას (დენატურაცია) გარემო პირობების შეცვლისას.
როდესაც ცილოვანი ხსნარები მჟავიანდება pH 4,5-5,0-მდე (მაგალითად, პიკინგის დროს), ცილები კარგავენ ხსნადობას და ნალექს (კოაგულაციას). ბევრი ცილა კარგავს ხსნადობას, როდესაც ხსნარი გაჯერებულია ნატრიუმის ქლორიდით (დამარილებით). კერძოდ, კუნთების ძირითადი პროტეინები, ძლიერ ხსნადი ნატრიუმის ქლორიდის ხსნარებში 7,5-10% კონცენტრაციით, ნალექი (დამარილებული), როდესაც მისი კონცენტრაცია იზრდება 15% -მდე. გაცხელებისას (ადუღების, შეწვის, გამოცხობის დროს) ცილები კოაგულაცია (კოაგულაცია) ხდება. ცილების თერმული დენატურაცია იწყება 28-35 o C-ზე. ცილების დენატურაცია ასევე ხდება მათი სისტემების დეჰიდრატაციის (გაუწყლოების) დროს (გაშრობის და გაყინვის დროს).
ცილების ნალექის (დამარილების, შედედების) დროს ირღვევა მათი კავშირი წყალთან.
სივრცითი სამგანზომილებიანი სტრუქტურის შედეგად ცილის მოლეკულის „ზედაპირზე“ არის ქიმიური აქტიური ჯგუფები– NN 2; -COOH; - ის. წყალხსნარში ეს ჯგუფები იონიზებულ მდგომარეობაშია სხვადასხვა ნიშნის მუხტით. ცილის მოლეკულა იძენს შესაბამისი ნიშანიდა მუხტის ოდენობა დამოკიდებულია დადებითად და უარყოფითად დამუხტული ჯგუფების თანაფარდობაზე. ცილის მოლეკულის მუხტი დამოკიდებულია მის მდგომარეობაზე. ცილის მოლეკულის სტრუქტურის ნებისმიერი ცვლილება იწვევს მისი მუხტის ცვლილებას; კერძოდ, მუხტის დაკარგვა იწვევს ცილის დენატურაციას. ამ მუხტების არსებობა ასევე განსაზღვრავს ცილების დამატენიანებელ თვისებებს. მაგალითად, წყლის მოლეკულები ემაგრება დადებითად დამუხტულ ცილის მოლეკულას მათი უარყოფითად დამუხტული ბოლოებით და იქმნება სტრუქტურა, რომლის ცენტრიც ცილის მოლეკულაა, ირგვლივ კი წყლის მონომოლეკულური გარსი. ვინაიდან წყლის მოლეკულების ყველა უარყოფითად დამუხტული ბოლო ცილის მოლეკულას აწყდება, იგივე მუხტი რჩება ცილა-წყლის სტრუქტურის ზედაპირზე. წყლის მოლეკულების ახალი ჯგუფები და ა.შ., თავის მხრივ, ერთვის ამ ზედაპირს. ამ შემთხვევაში, თითოეული ცილის მოლეკულის გარშემო წარმოიქმნება ელექტროსტატიკურად შეკრული დამატენიანებელი ფენა. პროტეინთან შეკავშირების სიძლიერე მცირდება ცენტრიდან მანძილის კვადრატის პროპორციულად, ე.ი. ცილის მოლეკულიდან და საკმარისად დიდ მანძილზე მოლეკულის ზომის მასშტაბით, ეს ბმა იმდენად მცირეა, რომ მოლეკულების საკუთარი თერმული მოძრაობა ხელს უშლის ელექტროსტატიკური ძალების მოქმედებას. ეს ზღუდავს ცილის ზედაპირზე შენახული წყლის რაოდენობას.
არსებული შეხედულებების მიხედვით, ცილოვანი ქსოვილი შეიძლება ჩაითვალოს ძალიან რთული სტრუქტურის კოლოიდური და კაპილარულ-ფოროვანი კოლოიდური სხეული, რომლის საფუძველია ადიდებულ მდგომარეობაში მყოფი ცილების სტრუქტურული ქსელი, რომელიც შეიცავს ბლანტი ხსნარებს, რომლებიც შეიცავს ხსნად ცილებს და სხვა აზოტებს. მინერალური ნივთიერებები, რომლებსაც აქვთ ჰიდროფილური თვისებები. ამავდროულად, წყლის ნაწილი შედის შემადგენლობაში კუნთების ქსოვილი, მყარად იჭერს სტრუქტურული ქსელის ცილებს, აგრეთვე გახსნილი ცილების და სხვა ჰიდროფილური ნივთიერებების მოლეკულებს.
ძალის ველის მიერ გამართულ წყალთან ერთად გარე და შიდა ზედაპირიპროტეინის ნაწილაკები, კუნთოვანი ქსოვილი შეიცავს წყალს, რომელიც შენარჩუნებულია ოსმოსურად და მექანიკური შემაკავშირებელი ძალებით (კაპილარული შეკავებული წყალი). ეს წყალი გვხვდება სითხეებში (ხსნარებში), რომლებიც შეიცავს სხვადასხვა აზოტოვან და ორგანულ ნივთიერებებს და მინერალურ მარილებს, ჩასმულია დახურულ უჯრედებში (მიკროპორებში) ცილის სტრუქტურებში და ამ უკანასკნელში შემავალი მიკრო და მაკროკაპილარებში. ლიტერატურაში არსებული მონაცემების მიხედვით, 1 გ ცილა ჰიდრატაციის დროს საშუალოდ 0,3 გ წყალს აკავშირებს.
დამუშავების ყველა მეთოდი და ტექნოლოგიური რეჟიმი მიზნად ისახავს ნედლეულის ქსოვილებში წყლის შეცვლას (მარილით გაჯერება, ყინულად გადაქცევა, დუღილის წერტილთან ახლოს ტემპერატურამდე გაცხელება, აორთქლება). შეცვლა შინაგანი ენერგიაწყალი იწვევს ცილასა და წყალს შორის წონასწორობის მდგომარეობის დარღვევას, რაც ქმნის დამატენიანებელ გარსს. ცილის მოლეკულა ამაზე რეაგირებს საკუთარი სტრუქტურის გადაწყობით და შესაბამისად მუხტის რაოდენობის შეცვლით. როდესაც ეს ცვლილებები იწვევს მუხტის მკვეთრ შემცირებას ან სრულ გაქრობას, ხდება ცილის დენატურაცია.
გარე გავლენის ინტენსივობიდან და ხანგრძლივობიდან გამომდინარე, ცილის დენატურაცია შეიძლება იყოს შექცევადი, ნაწილობრივ შექცევადი ან შეუქცევადი.
დენატურაციის სიღრმე შეიძლება განისაზღვროს კუნთოვანი ქსოვილის უნარით სრულად ან ნაწილობრივ აღადგინოს კავშირი წყალთან.
ამჟამად გამოყენებული მეთოდები საკვები ნედლეულის გადამუშავების მაღალი ცილის შემცველობით იწვევს ძირითადად ცვლილებებს, რომლებიც შეიძლება დახასიათდეს, როგორც ნაწილობრივი დენატურაცია. ცილის მოლეკულის დენატურაციის დიაგრამა ნაჩვენებია ნახ. 4.

ბრინჯი. 4. ცილის მოლეკულის დენატურაციის სქემა:
A - საწყისი მდგომარეობა, B - საწყისი შექცევადი განვითარება, C - მოწინავე შეუქცევადი განვითარება.
ცილის ყველაზე დამახასიათებელი ცვლილებები თერმული დენატურაციის დროს (ტემპერატურა 70-100°C) არის მისი ბუნებრივი თვისებების დაკარგვა (წყალში დაშლის უნარი, მარილების და სპირტების ხსნარი), ასევე შეშუპების უნარის დაქვეითება. .
თერმულ დენატურაციასთან დაკავშირებული პროტეინის ცვლილებები უფრო მნიშვნელოვანია, რაც უფრო მაღალია გათბობის ტემპერატურა და ხანგრძლივობა, წნევის მოქმედება და წყალხსნარში ცილა უფრო სწრაფად დენატურდება, ვიდრე გამხმარ მდგომარეობაში.
ცილის დენატურაცია მნიშვნელოვან როლს ასრულებს რიგ ტექნოლოგიური პროცესები: პურის, საკონდიტრო ნაწარმის გამოცხობისას, ხორცის, თევზის, ბოსტნეულის, რძისა და კვერცხის ფხვნილის გაშრობისას, კონსერვის დროს და ა.შ.
როდესაც პროდუქტი სრულ მზადყოფნაში მიდის, ჩვეულებრივ, 100°C-მდე ტემპერატურაზე ხანგრძლივი ზემოქმედებით, ცილები განიცდიან შემდგომ ცვლილებებს, რომლებიც დაკავშირებულია მათი მაკრომოლეკულების განადგურებასთან - ჰიდროლიზით.
პროცესის დასაწყისში, აქროლადი პროდუქტები შეიძლება გამოიყოს ცილის მოლეკულებიდან: ნახშირორჟანგიწყალბადის სულფიდი, ამიაკი, წყალბადის ფოსფორი და სხვა ნივთიერებები, რომლებიც მონაწილეობენ მზა პროდუქტების გემოსა და არომატის ფორმირებაში. წყლისა და სითბოს გახანგრძლივებული ზემოქმედებით, წყალში ხსნადი აზოტოვანი ნივთიერებების წარმოქმნა ხდება ცილის მოლეკულის დეპოლიმერიზაციის გამო, რაც ხდება, მაგალითად, კოლაგენის გლუტინზე გადასვლის დროს.
ცილის ჰიდროლიზი შეიძლება გამოწვეული იყოს პროტეოლიზური ფერმენტების დახმარებით, რომლებიც გამოიყენება გარკვეული ტექნოლოგიური პროცესების გასაძლიერებლად (მყარი ხორცის დარბილება, საფუარის ცომის მომზადება და ა.შ.).
ბიოქიმიის ინტერნეტი
ბიოქიმია
შემდეგი განყოფილებაცილები. ამინომჟავები -- ცილების სტრუქტურული კომპონენტები
პროტეინები არის აზოტის შემცველი, მაღალმოლეკულური ორგანული ნაერთები, რომლებიც შედგება ამინომჟავებისგან, რომლებიც დაკავშირებულია ჯაჭვებში პეპტიდური ბმების გამოყენებით და აქვთ რთული სტრუქტურული ორგანიზაცია.
იგივე ამინომჟავები გვხვდება სხვადასხვა სტრუქტურისა და ფუნქციის მქონე ცილებში. ცილის მოლეკულების ინდივიდუალობა განისაზღვრება ცილაში ამინომჟავების მონაცვლეობის თანმიმდევრობით.
დამახასიათებელი ნიშნებიცილები, რომლებიც განასხვავებენ მათ უჯრედის სხვა ორგანული ნაერთებისგან:
1. ცილები აზოტის შემცველი ნაერთებია, ისევე როგორც მრავალი სხვა უჯრედის კომპონენტი (ნუკლეინის მჟავები, ზოგიერთი ლიპიდი, ნახშირწყლები), მაგრამ სხვა ორგანული ნივთიერებებისგან განსხვავებით, აზოტის შემცველობა გაცილებით მაღალია - საშუალოდ 16 გრამი 100 გრამ ცილაზე.
2. ცილების სტრუქტურული ერთეულია L-სერიის ალფა ამინომჟავები.
3. ამინომჟავები დაკავშირებულია ცილებში პეპტიდური ბმების გამოყენებით, ქმნიან პოლიპეპტიდურ ჯაჭვს.
4.პროტეინებს აქვთ დიდი მოლეკულური წონა (20000-დან რამდენიმე მილიონ დალტონამდე).
5. ცილები განსხვავდებიან რთული სტრუქტურული ორგანიზაციით (მათ აქვთ პირველადი, მეორადი, მესამეული და მეოთხეული სტრუქტურა).
ცილების მრავალფეროვანი ფუნქციები:
1.კატალიტიკური. ბევრი ცილა არის ფერმენტი
2. მარეგულირებელი. ზოგიერთი ჰორმონი არის ცილა
3.სტრუქტურული. ცოცხალი უჯრედის ყველა სტრუქტურა შეიცავს ცილებს.
4. რეცეპტორი. ცილები რეცეპტორების აუცილებელი კომპონენტია და შეუძლიათ სხვა მოლეკულების ამოცნობა.
5. ტრანსპორტი. ისინი გადააქვთ ცხიმებს, პიგმენტებს, ჰორმონებს, წამლებს, სხვადასხვა ჰიდროფობიურ ნაერთებს და ა.შ.
6. მხარდაჭერა. კოლაგენი, ძვლის ქსოვილის ცილები.
7.ენერგია. 1 გ ცილის დაჟანგვას თან ახლავს 17,6 კჯ ენერგიის გამოყოფა. ენერგიის 15% დღეში წარმოიქმნება ცილების დაშლის გამო.
8.კონტრაქტული. ამ ფუნქციას ასრულებენ კუნთოვანი ქსოვილის პროტეინები - აქტინი, მიოზინი.
9.ჰისტონის გენის მარეგულირებელი ფუნქცია.
10.იმუნოლოგიური. ანტისხეულები არის ცილები.
11. ჰემოსტატიკური. ფიბრინოგენი და სისხლის პლაზმის სხვა ცილები მონაწილეობენ სისხლის შედედების პროცესებში.
1.1.ამინომჟავები -- ცილების სტრუქტურული კომპონენტები
ამინომჟავები ჰეტეროფუნქციური ნაერთებია. ისინი წარმოადგენენ კარბოქსილის მჟავების წარმოებულებს, რომლებშიც წყალბადის ატომი ალფა მდგომარეობაში ჩანაცვლებულია ამინო ჯგუფით.
ყველა ამინომჟავა, რომელიც ქმნის ცილებს, ხასიათდება შემდეგი მახასიათებლებით:
ა) ამინო ჯგუფის პოზიციის მიხედვით ისინი ა-ამინომჟავებია (ამინო ჯგუფი კარბოქსილის ჯგუფის გვერდით არის).
ბ) ისინი ყველა მიეკუთვნება L-სერიას, ვინაიდან ასიმეტრიული (ქირალური) ნახშირბადის ატომის ამინო ჯგუფი მარცხნივ იწერება. გამონაკლისი არის გლიცინი, რომელსაც არ აქვს ასიმეტრიული ნახშირბადის ატომი; მისი გვერდითი ჯაჭვი წარმოდგენილია წყალბადის ატომით.
გ) აქვთ ერთი და იგივე ზოგადი ფრაგმენტი და განსხვავდებიან მხოლოდ გვერდითი ჯაჭვის აგებულებით (R), რომლის თვისებები დიდწილად განსაზღვრავს თავად ამინომჟავების თვისებებს და იმ ცილებს, რომლებშიც ისინი შედის. ეს არის განსხვავებები ფორმაში, ზომასა და პოლარობაში, რაც საშუალებას აძლევს ამინომჟავებს იყოს სამშენებლო ბლოკები, რომელსაც ევოლუცია იყენებს ცილის სტრუქტურის მკაცრი მოთხოვნების დასაკმაყოფილებლად.
დ) ხსნარებში ნეიტრალური pH მნიშვნელობებით, ალფა ამინომჟავები ძირითადად გვხვდება შიდა მარილების სახით. ძლიერ მჟავე გარემოში ჭარბობს კათიონური ფორმა, ძლიერ ტუტე გარემოში – ანიონური ფორმა. (
მრავალი ამინომჟავა, რომელიც არ არის ცილების ნაწილი (ე.წ. არაცილოვანი ამინომჟავები) აღმოჩენილია სხვადასხვა ორგანიზმში. თუმცა, ყველა ცნობილი ორგანიზმი იყენებს ერთსა და იმავე 20 ამინომჟავას მათი ცილების შესაქმნელად; ფ. კრიკმა მათ "ჯადოსნური ოცი" უწოდა. მხოლოდ ისინი დაშიფრულია გენეტიკური კოდით.
ამინომჟავების სტრუქტურა და კლასიფიკაცია
არსებობს სხვადასხვა კლასიფიკაციაამინომჟავები, რომლებიც ქმნიან ცილებს, იმისდა მიხედვით, თუ რა მახასიათებელია მათი ჯგუფებად დაყოფის საფუძველი. ასე რომ, გვერდითი ჯაჭვის ქიმიური ბუნების მიხედვით, ა-ამინომჟავები იყოფა ალიფატურ (აციკლურ), არომატულ და ჰეტეროციკლურებად.
I. Aliphatica - ამინომჟავები
1. მონოამინომონოკარბოქსილის (ნეიტრალური)
ალიფატური ამინომჟავები, რომლებიც შეიცავს დამატებით ფუნქციურ ჯგუფს გვერდით ჯაჭვში.
ა) ჰიდროქსილის ჯგუფი (ჰიდროქსიამინომჟავები)
ბ) კარბოქსილის ჯგუფი (მონოამინოდიკარბოქსილი)
ბ) ამიდური ჯგუფი
დ) ამინოჯგუფი (დიამინომონოკარბოქსილი)
II. არომატული ამინომჟავები III. ჰეტეროციკლური ამინომჟავები
IV. იმინომჟავა
ჰიდროფილური (პოლარული) ჯგუფებია ჯგუფები: -OH, -SH, -COOH, - NH2, იმიდაზოლის ბირთვი. ჰიდროფილურობა არის მოლეკულების ან ჯგუფების თვისება, რომლებსაც აქვთ... ამასთან დაკავშირებით 20-ვე ამინომჟავა შეიძლება დაიყოს...არაპოლარული ჰიდროფობიური,
პოლარული არაიონური,
პოლარული უარყოფითად დამუხტული,
პოლარული დადებითად დამუხტულია.
ამ ამინომჟავების გვერდითი ჯაჭვი შეიცავს არაპოლარულ, არაიონურ ჯგუფებს. ამ კლასში შედის ალიფატური ამინომჟავები. გლიცინი არის ყველაზე... პოლარული, არაიონური R-ჯგუფების შემცველი ამინომჟავები.ამ ჯგუფის ამინომჟავები შეიცავს არაიონურ ჯგუფებს გვერდით ჯაჭვში, რომლებსაც არ შეუძლიათ პროტონის დონაცია ან მიღება. ამ ამინომჟავების გვერდითი ჯგუფები წყალში იხსნება, რადგან ისინი შეიცავენ პოლარულ ფუნქციურ ჯგუფებს, რომლებიც ქმნიან წყალბადურ კავშირებს წყალთან. მათ შორისაა სერინი, თრეონინი, რომლებიც შეიცავს სპირტის ჰიდროქსილს გვერდით ჯაჭვში, ასევე გლუტამინს, ასპარაგინს, რომლებიც შეიცავს ამიდურ ჯგუფებს. ცისტეინი და ტიროზინი ასევე მიეკუთვნება ამ ჯგუფს. ეს ამინომჟავები შეიცავს, შესაბამისად, თიოლის ჯგუფს და ფენოლურ ჰიდროქსილს, რომლებსაც შეუძლიათ დისოციაცია, მაგრამ უჯრედებში შენარჩუნებული ნეიტრალური pH მნიშვნელობებით, ეს ჯგუფები პრაქტიკულად არ იშლება.
ამინომჟავები პოლარული უარყოფითი მუხტით
R- ჯგუფები.
ამ ჯგუფში შედის ასპარტინული და გლუტამინის ამინომჟავები, რომლებსაც აქვთ დამატებითი კარბოქსილის ჯგუფი გვერდით ჯაჭვში, რომელსაც შეუძლია დისოციაცია.
ამინომჟავები პოლარული, დადებითად დამუხტული R-ჯგუფებით. ლიზინს აქვს დამატებითი დადებითად დამუხტული ჯგუფი გვერდითა ჯაჭვში,... წინა განყოფილება ზედა დონის სექცია შემდეგი განყოფილებაცილის მოლეკულების სტრუქტურული ორგანიზაციის დონეები
ცილის მოლეკულების სტრუქტურა ხასიათდება მნიშვნელოვანი სირთულით და უნიკალური ორგანიზებით. ცილის სტრუქტურული ორგანიზაციის 4 დონეა:... პირველადი სტრუქტურა არის ამინომჟავების თანმიმდევრობა პოლიპეპტიდურ ჯაჭვში,... ამინომჟავების თანმიმდევრობა პოლიპეპტიდურ ჯაჭვში განსაზღვრავს ცილის სტრუქტურული ორგანიზაციის შემდგომ დონეებს, მისი...პროტეინის მოდულები (დომენი)
ცხიმოვანი მჟავა სინთაზას, რომელიც წარმოადგენს ერთ პოლიპეპტიდურ ჯაჭვს, აქვს 7 დომენი 7 რეაქციის კატალიზებისთვის. ვარაუდობენ, რომ სინთაზას დომენები ერთხელ...ცილის აქტიური ცენტრი და მისი ურთიერთქმედება ლიგანდთან.
ცილების ფუნქციონირება ეფუძნება მათ სპეციფიკურ ურთიერთქმედებას ლიგანდებთან. უნიკალური აქტიური შემცველი 50000 ინდივიდუალური ცილა... მეოთხეული სტრუქტურა არის უმაღლესი დონესტრუქტურული ორგანიზაცია, შესაძლებელია... მეოთხეული სტრუქტურა სტაბილიზირებულია არაკოვალენტური ბმებით, რომლებიც წარმოიქმნება პროტომერების კონტაქტურ უბნებს შორის,...ცილების ფიზიკოქიმიური თვისებები
ცილების პირველადი სტრუქტურა დიდწილად განსაზღვრავს მეოთხეული სტრუქტურის მეორად, მესამეულ სტრუქტურებსა და თავისებურებებს. თავის მხრივ,... ცილების მოლეკულური წონა საკმაოდ დიდია, ამიტომ ისინი მიეკუთვნებიან... ზოგიერთი ცილის მოლეკულური წონაა: ინსულინი - 5700D,ცილების კლასიფიკაცია
ადამიანის სხეული შეიცავს 50000-ზე მეტ ინდივიდუალურ ცილას, რომლებიც განსხვავდებიან პირველადი სტრუქტურით, კონფორმაციით, აქტიური ცენტრის სტრუქტურით და ფუნქციებით. თუმცა, დღემდე არ არსებობს ერთიანი და ჰარმონიული კლასიფიკაცია, რომელიც ითვალისწინებს სხვადასხვა თვისებებიცილები. არსებული კლასიფიკაცია ეფუძნება სხვადასხვა მახასიათებლებს. ასე რომ, ცილები შეიძლება დაიყოს:
· ცილის მოლეკულების ფორმის მიხედვით (გლობულური - მრგვალი ან ბოჭკოვანი - ძაფისებრი)
· მოლეკულური წონის მიხედვით (დაბალი მოლეკულური წონა, მაღალი მოლეკულური წონა)
· შესრულებული ფუნქციებით (სატრანსპორტო, სტრუქტურული, დამცავი, მარეგულირებელი და ა.შ.)
· უჯრედში ლოკალიზაციით (ბირთვული, ციტოპლაზმური, ლიზოსომური და ა.შ.)
· სტრუქტურული მახასიათებლების მიხედვით და ქიმიური შემადგენლობაცილები იყოფა ორ ჯგუფად: მარტივი და რთული. მარტივი ცილები წარმოდგენილია მხოლოდ პოლიპეპტიდური ჯაჭვით, რომელიც შედგება ამინომჟავებისგან. კომპლექსურ პროტეინებს აქვთ ცილოვანი ნაწილი და არაცილოვანი კომპონენტი (პროთეზური ჯგუფი). თუმცა, ეს კლასიფიკაცია არ არის იდეალური, რადგან სუფთა ფორმამარტივი ცილები ორგანიზმში იშვიათია.
მარტივი ცილების მახასიათებლები.
ჰისტონები არის მრავალი ორგანიზმის ქსოვილის ცილები, რომლებიც დაკავშირებულია ქრომატინის დნმ-თან. ეს არის მცირე მოლეკულური წონის ცილები (11-24 ათასი და). მიხედვით... არსებობს ჰისტონების 5 სახეობა. დაყოფა ემყარება მთელ რიგ მახასიათებლებს, რომელთაგან მთავარია... ჰისტონების ძირითადი ფუნქცია სტრუქტურული და მარეგულირებელია. სტრუქტურული ფუნქცია არის ის, რომ ჰისტონები მონაწილეობენ ...ალბუმინები და გლობულინები.
A და G ცილები, რომლებიც გვხვდება ყველა ქსოვილში. ამ ცილებით ყველაზე მდიდარი სისხლის შრატია. მასში ალბუმინის შემცველობა არის 40-45 გ/ლ,... ალბუმინები შედარებით დაბალი მოლეკულური წონის ცილებია (15-70 ათასი დიახ); ისინი... მაღალი ჰიდროფილურობის, მცირე მოლეკულური ზომების და მნიშვნელოვანი კონცენტრაციის გამო, ალბუმინები მნიშვნელოვან როლს თამაშობენ...პროლამინი და გლუტელინები.
გლუტელინები ასევე მცენარეული ცილებია, რომლებიც არ იხსნება წყალში, მარილის ხსნარებში და ეთანოლში. ისინი ხსნადია სუსტ ტუტეებში.პროტეინოიდები.
ყველა ეს ცილა ფიბრილარულია და არ ჰიდროლიზდება კუჭ-ნაწლავის ტრაქტში. ნაწლავის ტრაქტი. კოლაგენი შეადგენს მთლიანი ცილის 25-33%-ს... ელასტინი არის ელასტიური ბოჭკოების მთავარი სტრუქტურული კომპონენტი, რომელიც...რთული ცილები
კომპლექსური ცილები, პოლიპეპტიდური ჯაჭვების გარდა, შეიცავს არაცილოვან (პროთეზულ) ნაწილს, რომელიც წარმოდგენილია სხვადასხვა ნივთიერებებით. არაცილოვანი ნაწილის ქიმიური ბუნებიდან გამომდინარე, ისინი იყოფა შემდეგი ჯგუფებირთული ცილები:
· ქრომოპროტეინები
· ნახშირწყლო-ცილოვანი კომპლექსები
· ლიპიდურ-ცილოვანი კომპლექსები
· ნუკლეოპროტეინები
· ფოსფოპროტეინები
ქრომოპროტეინები
ქრომოპროტეინებირთული პროტეინებია, რომელთა პროთეზური ნაწილი წარმოდგენილია ფერადი კომპონენტით (ბერძნულიდან Chromos - საღებავი). მათ შორისაა ბიოლოგიურად მნიშვნელოვანი ცილები ჰემოგლობინი, მიოგლობინი, ასევე ზოგიერთი ფერმენტი: კატალაზა, პეროქსიდაზა, ციტოქრომები, ყველა მათგანი ჰემეპროტეინებია, რადგან მათი პროთეზირების ნაწილი შეიცავს ჰემს.
ჰემოგლობინი (Hb).
Hb ქვედანაყოფები ერთმანეთს „ამოიცნობენ“ მათ ზედაპირზე იმ რეგიონების არსებობის გამო, რომლებიც ავსებენ ფორმას. თითოეული ქვედანაყოფი ან პოლიპეპტიდი... ჰემოგლობინის თითოეული პროტომერი ბუნებრივი კოორდინაციაა...ჰემოგლობინის სახეები.
ჰემოგლობინის ფიზიოლოგიური ტიპები განსხვავდება ერთმანეთისგან პოლიპეპტიდური ჯაჭვების ან ქვედანაყოფების სიმრავლით, რომლებიც წარმოიქმნება სხვადასხვა ეტაპებიგანვითარება... ა) პრიმიტიული NBP, ყველაზე მეტად ჩნდება ადრეული ეტაპებიემბრიონის განვითარება (1 –… ბ) ნაყოფის ჰემოგლობინი HbF (ლათ. Fetus - fetus). HbF არის ნაყოფის ჰემოგლობინის ძირითადი ტიპი და წარმოადგენს...გლიკოპროტეინები (GP)
პროტეოგლიკანები
GP-ის მოკლე ნახშირწყლების ჯაჭვები აგებულია გლუკოზამინის, გალაქტოზამინის, გლუკოზისა და გალაქტოზისგან. GP-ს შემადგენლობაში ყველაზე მნიშვნელოვანი მონოსაქარიდებია N –... ნახშირწყლების მოკლე ჯაჭვების რაოდენობა GP-ში შეიძლება 300-800-ს მიაღწიოს. სიგრძე და...მაღალი სპეციფიკური ამოცნობის შერჩევითი ურთიერთქმედების ფუნქცია.
მემბრანების ზედაპირზე განლაგებული უჯრედული GP მონაწილეობს ბიოლოგიური ამოცნობისა და უჯრედშორისი ურთიერთქმედების ძალიან დახვეწილ პროცესებში, მოქმედებენ როგორც რეცეპტორული სისტემები გარკვეული ნაერთებისა და უჯრედებისთვის.
ტრანსპორტის როლი.
ჰიდროფობიური ნივთიერებებისა და ლითონის იონების ტრანსპორტირებას GP-ები. Ისე
რკინის მატარებლის ფუნქციას ასრულებს GP – ტრანსფერინი; სპილენძი – ცერულოპლაზმინი; სტეროიდული ჰორმონები - ტრანსკორტინი.
კატალიზური.
ნახშირწყლების კომპონენტი გვხვდება ზოგიერთ ფერმენტში: ენტეროკინაზა, პეროქსიდაზა, გლუკოზის ოქსიდაზა, ქოლინესტერაზა.
დამცავი შეზეთვის ფუნქცია.
გლიკოპროტეინები არის სანერწყვე მუცინების, კუჭისა და ნაწლავების მუცინების კომპონენტები.
მონაწილეობა მიიღოს სისხლის შედედების პროცესში.
პროტეოგლიკანები. ეს არის ნახშირწყლო-ცილოვანი კომპლექსები, რომელთა ნახშირწყლების კომპონენტი წარმოდგენილია ჰეტეროპოლისაქარიდებით, აგებული...უფასო ლიპოპროტეინები.
სისხლის პლაზმისა და რძის ლიპოპროტეინები წყალში ხსნადია.
სტრუქტურული პროტეოლიპიდები.
ისინი ბიომემბრანების ნაწილია და ცხიმში ხსნადია.
ლიპიდური - ცილის კომპლექსები შეიცავს ლიპიდურ კომპონენტებს, როგორც არაცილოვან ნაწილს.
უმაღლესი ცხიმოვანი მჟავები
უფასო ლიპოპროტეინები.
LA ფრაქციები: ა) ქილომიკრონები (CM). ეს არის ყველაზე დაბალი სიმკვრივის ფრაქცია, რადგან შემადგენლობაში... ბ) ძალიან დაბალი სიმკვრივის ლიპოპროტეინები (VLDL).ან პრე-ბ-ლიპოპროტეინები მათი სიმკვრივეა 0,94 - 1,006 კგ/ლ;სტრუქტურული ლიპოპროტეინები (პროტეოლიპიდები).
ნუკლეოპროტეინები
ნუკლეინის მჟავები არის მაღალი მოლეკულური წონის ნაერთები, რომლებიც შედგება მონონუკლეოტიდებისგან, ე.ი. მათი სტრუქტურული ერთეულია მონონუკლეოტიდი... ფუძისა და პენტოზის შეერთებას ნუკლეოზიდი ეწოდება, ბმა პენტოზასა და აზოტოვან ფუძეს შორის (b - გლიკოზიდური)...ყველაზე გავრცელებული ნუკლეოტიდების ნომენკლატურა.
1. ადენოზინის მონოფოსფატი (AMP), ადენილის მჟავა. 2. გუანოზინის მონოფოსფატი (GMP), გუანილის მჟავა. 3. ციტიდინ მონოფოსფატი (CMP), ციტიდილის მჟავა.ნუკლეინის მჟავების სტრუქტურა.
ამ შემთხვევაში ეთერულ ბმას წარმოქმნის ერთი მონონუკლეოტიდის ფოსფატური ნარჩენი და მეორის პენტოზის ნარჩენების 3" ჰიდროქსილის ჯგუფი... პოლინუკლეოტიდების ბოლოები განსხვავდება აგებულებით: ერთ ბოლოში არის...დნმ-ის მეორადი სტრუქტურა.
ამ მოდელის მიხედვით, დნმ-ის მოლეკულა არის ორმაგი სპირალი, რომელიც წარმოიქმნება ერთმანეთის მიმართ გადაბმული ორი პოლინუკლეოტიდური ჯაჭვისგან... დნმ-ის ჯაჭვების ყველა ფუძე (ჰიდროფობიური თვისებებით) განლაგებულია ორმაგში... წყალბადური ბმების გარდა. ჰიდროფობიური ურთიერთქმედება, რომელიც წარმოიქმნება დროს...რნმ-ის სტრუქტურის თავისებურებები.
რნმ-ის პირველადი სტრუქტურამსგავსია დნმ-ის პირველადი სტრუქტურისა და წარმოადგენს პოლინუკლეოტიდურ ჯაჭვს, რომელიც შედგება მონონუკლეოტიდებისგან, რომლებიც დაკავშირებულია 3" და 5" ფოსფოდიესტერული ბმებით.
რნმ-ის მეორადი სტრუქტურა.
რნმ-ის ძირითადი ტიპები.
1. გადაცემის რნმ (tRNAs) tRNA-ების სივრცითი სტრუქტურა, განსხვავებების მიუხედავად თანმიმდევრობით...ნუკლეინის მჟავების მესამეული სტრუქტურა.
მიტოქონდრიისა და ქლოროპლასტების ზოგიერთი დნმ-ის ვირუსის შესწავლამ ფიზიკური, ფიზიკოქიმიური მეთოდების გამოყენებით აჩვენა, რომ დნმ-ის ორმაგი სპირალი... სუპერსპირალი სტრუქტურა (სუპერკულული) უზრუნველყოფს ეკონომიურ შეფუთვას... tRNA-ს სტრუქტურის შესახებ არსებული მონაცემები მიუთითებს, რომ ადგილობრივი tRNA მოლეკულები დაახლოებით იგივე აქვს...ფოსფოპროტეინები.
დადგენილია, რომ უჯრედებში ფოსფოპროტეინები სინთეზირდება ფოსფორილირების შედეგად პროტეინ კინაზების მონაწილეობით. ფოსფოპროტეინებში შედის რძის კაზეინოგენი, რომელიც არის ცილა...ნახშირწყლები. ნახშირწყლების კლასიფიკაცია
მე-19 საუკუნეში შემოთავაზებული ტერმინი „ნახშირწყლები“ ემყარებოდა იმ ვარაუდს, რომ ყველა ნახშირწყალი შეიცავს სამ ელემენტს - ნახშირბადს, წყალბადს და ჟანგბადს, ამ უკანასკნელის თანაფარდობა წყალშია და ელემენტარული შემადგენლობა შეიძლება გამოიხატოს ფორმულით. Cn (H 2 O)m. თუმცა, როგორც ახალი ნახშირწყლები აღმოაჩინეს, მათ აღმოაჩინეს, რომ ყველა მათგანი არ აკმაყოფილებდა ამ ფორმულას.
ტერმინი „ნახშირწყლები“ მოძველებულია და არ ასახავს ამ ნაერთების არც ქიმიურ ბუნებას და არც შემადგენლობას, თუმცა მათთვის შემოთავაზებული ტერმინი „გლიციდები“ ფართოდ არ გავრცელებულა. ნახშირწყლების დამახასიათებელი განმასხვავებელი ნიშანია მათ შემადგენლობაში არსებობა. მინიმუმ ორი ჰიდროქსილის ჯგუფი და კარბონილის (ალდეჰიდის ან კეტონის) ჯგუფი. ჯგუფები, ანუ ნახშირწყლები არის პოლიოქსიკარბონილის ნაერთები და მათი წარმოებულები.
ნახშირწყლები ბუნებაში ორგანული ნაერთების ყველაზე გავრცელებული კლასია. უჯრედებში ნახშირწყლების ფუნქციები ძალიან მრავალფეროვანია. ისინი ემსახურებიან როგორც უჯრედის ენერგიის წყაროს და აკუმულატორს, ასრულებენ სტრუქტურულ როლს; ისინი, გლიკოზამინოგლიკანების სახით, უჯრედშორისი მატრიქსის ნაწილია და მონაწილეობენ ბევრ მეტაბოლურ პროცესში.
მონოსაქარიდები არის ნახშირწყლები, რომლებიც არ განიცდიან ჰიდროლიზს, ე.ი. არ დაიშალოს უფრო მარტივ შაქარებად. ოლიგოსაქარიდები - რთული ნახშირწყლები, რომელიც შეიცავს 2-დან 10-მდე მონოსაქარიდის ნარჩენებს.
პოლისაქარიდები არის მაღალმოლეკულური ნაერთები, რომელთა მაკრომოლეკულები შეიცავს ასობით და ათასობით მონოსაქარიდის ნარჩენებს ერთი ტიპის (ჰომოპოლისაქარიდები) ან განსხვავებული ტიპები(ჰეტეროპოლისაქარიდები).
2.2.მონოსაქარიდები
მონოსაქარიდების კლასიფიკაცია ემყარება ორ მახასიათებელს:
ფუნქციური ჯგუფების ხელმისაწვდომობა.
ნახშირბადის ატომების რაოდენობა მონოსაქარიდებში.
მოლეკულაში კარბონილის ჯგუფის პოზიციიდან გამომდინარე, მონოსაქარიდები იყოფა ალდოზებად და კეტოზებად. ალდოზები შეიცავს ალდეჰიდის ჯგუფს, ხოლო კეტოზები შეიცავს კეტო ჯგუფს.
ნახშირბადის ატომების რაოდენობის მიხედვით განასხვავებენ მონოსაქარიდების შემდეგ ჯგუფებს: ტრიოზები, ტეტროზები, პენტოზები, ჰექსოზები, ჰეპტოზები.
მონოსაქარიდების ღია (დახურული) ფორმები გამოსახულია ფიშერის პროექციის ფორმულების სახით. მონოსაქარიდის მოლეკულები შეიცავს რამდენიმე ქირალურ ცენტრს (C*) და მიეკუთვნება L- ან D-სერიის ნაერთებს.
მონოსაქარიდების შედარებითი კონფიგურაცია განისაზღვრება კონფიგურაციის სტანდარტით - გლიცერალდეჰიდი.
მონოსაქარიდები მიეკუთვნებიან D-სერიებს, თუ OH ჯგუფი ქვედა ქირალურ ნახშირბადის ატომში არის მარჯვნივ და L-სერიას, თუ OH ჯგუფი მარცხნივ არის. ამიტომ, ქვედა ქირალურ ატომში მდებარე ჰიდროქსილს ეწოდება D-, L-განმსაზღვრელი.
წყალხსნარში არის მონოსაქარიდების როგორც ღია, ასევე ციკლური ფორმები. ციკლური ფორმებია ხუთწევრიანი (ფურანოზი) და ექვსწევრიანი (პირანოზა) ჰეტეროციკლები.
ციკლების სახელები მომდინარეობს მონათესავე ნაერთების სახელებიდან - ფურანი და პირანი
მონოსაქარიდების ციკლური ფორმებია ჰემიაცეტალები. ისინი წარმოიქმნება მონოსაქარიდის კარბონილისა და ჰიდროქსილის ჯგუფებს შორის ინტრამოლეკულური ურთიერთქმედების შედეგად. გვხვდება ციკლური ფორმით ახალი ცენტრიქირალობა, ასევე ახალი ჰიდროქსილი - ჰემიოციტალი. ეს იწვევს იზომერების კიდევ ერთი წყვილის წარმოქმნას, რომელსაც ანომერებს უწოდებენ.
მონოსაქარიდების ციკლური ფორმებისთვის მიღებულია ჰაუორტის პერსპექტიული ფორმულები, რომლებშიც ციკლები გამოსახულია როგორც ბრტყელი მრავალკუთხედები, რომლებიც მდებარეობენ ნახატის სიბრტყის პერპენდიკულარულად. ჟანგბადის ატომი მდებარეობს პირანოზის რგოლის უკიდურეს მარჯვენა კუთხეში, ხოლო ბეჭდის სიბრტყის უკან ფურანოზის რგოლში. ციკლებში ნახშირბადის ატომების სიმბოლოები არ იწერება.
ფიშერის პროექციის ფორმულებიდან Haworth-ის ფორმულებზე გადასასვლელად, თქვენ უნდა გაითვალისწინოთ შემდეგი: ატომები და ატომების ჯგუფები, რომლებიც მდებარეობს ფიშერის ფორმულებში ნახშირბადის ჯაჭვის მარცხნივ, Haworth ფორმულებში განლაგებულია ციკლის სიბრტყის ზემოთ; შემცვლელები, რომლებიც მდებარეობს მარჯვნივ - თვითმფრინავის ქვეშ.
ზემოაღნიშნულის გათვალისწინებით, Haworth-ის ფორმულებში ყველაზე მნიშვნელოვან მონოსაქარიდებს აქვთ შემდეგი ფორმა.
მონოსაქარიდის წარმოებულები
სიალიუმის მჟავები დიდწილად განსაზღვრავს ლიგანდის ურთიერთქმედებას რეცეპტორებთან...ოლიგოსაქარიდები
ლაქტოზა და მალტოზა ამცირებს დისაქარიდებს. მათში გლიკოზიდური ბმა იქმნება ჰემიაცეტალური (გლიკოზიდური) OH ჯგუფის გამო... მალტოზასა და ლაქტოზასგან განსხვავებით, საქაროზა არააღმდგენია... მალტოზა (ალაოს შაქარი) წარმოიქმნება ნაწლავებში სახამებლის დაშლის დროს. შეიცავს დიდი რაოდენობითალაოსში...ჰომოპოლისაქარიდები (PS).
- მონოსაქარიდების სტრუქტურა, რომლებიც ქმნიან ჯაჭვს - გლიკოზიდური ბმების ტიპი, რომელიც აკავშირებს მონომერებს ჯაჭვში - მონოსაქარიდის ნარჩენების თანმიმდევრობა ჯაჭვში.ჰეტეროპოლისაქარიდები
გლიკოზამინოგლიკანები არის ჰეტეროპოლისაქარიდების გრძელი განშტოებული ჯაჭვები, აგებული განმეორებადი დისაქარიდის ერთეულებისგან - დიმერებისგან... ძირითადი გლიკოზამინოგლიკანებია: ჰიალურონის მჟავა,... ჰიალურონის მჟავა აგებულია განმეორებადი ერთეულებისგან, მათ შორის გლუკურონის მჟავა და N - აცეტილგლუკოზამინი.ლიპიდები
ლიპიდები
ტერმინი "ლიპიდები" აერთიანებს ნივთიერებებს, რომლებსაც აქვთ საერთო ფიზიკური თვისება - ჰიდროფობიურობა, ე.ი. წყალში უხსნადობა. სტრუქტურულად, ლიპიდები სხვადასხვა ქიმიური სტრუქტურის ნაერთებია. ისინი იყოფა კლასებად, რომლებიც აერთიანებენ მოლეკულებს, რომლებსაც აქვთ მსგავსი ქიმიური სტრუქტურა და საერთო ბიოლოგიური თვისებები.
ორგანიზმში ლიპიდების ძირითადი ნაწილია ცხიმები- ტრიაცილგლიცეროლები, რომლებიც ენერგიის შესანახი ფორმაა.
ფოსფოლიპიდები არის ლიპიდების დიდი კლასი, რომელიც შეიცავს ფოსფორის მჟავას ნარჩენებს, რაც მათ ამფიფილურ თვისებებს ანიჭებს. ამ თვისების გამო, ფოსფოლიპიდები ქმნიან ორშრიანი მემბრანულ სტრუქტურას, რომელშიც ჩაეფლო ცილები.
სტეროიდები, რომლებიც წარმოდგენილია ცხოველთა სამყაროში ქოლესტერინით და მისი წარმოებულებით, ასრულებენ სხვადასხვა ფუნქციებს.
ცხიმოვანი მჟავები და აცილგლიცეროლები.
ცხიმოვანი მჟავები სხვადასხვა ლიპიდების სტრუქტურული კომპონენტებია. ცხიმოვანი მჟავები ადამიანის ლიპიდებში არის სწორი ჯაჭვის ნახშირწყალბადები... ადამიანის ორგანიზმში ცხიმოვან მჟავებში ორმაგი ბმები ცის კონფიგურაციაშია. ...გაჯერებული მჟავებიფოსფოლიპიდები და სფინგოლიპიდები
ფოსფოლიპიდები
გლიცეროფოსფოლიპიდები გლიცეროლის წარმოებულებია, რომლებშიც ნახშირბადის პირველი და მეორე ატომები დაკავშირებულია ესტერული ბმებით ცხიმოვანი მჟავების ნარჩენებთან... მცირე რაოდენობით, მაგრამ არის შუალედური პროდუქტი სინთეზის გზაზე... გლიცეროფოსფოლიპიდებში, ნახშირბადის პირველი ატომი. გლიცერინი ესტერიფიცირებულია გაჯერებული ცხიმოვანი მჟავებით, მეორე...სფინგოლიპიდები
კერამიდებში სფინგოზინის სპირტი არაჩვეულებრივად ასოცირდება ცხიმოვან მჟავებთან (ამიდური ბმა), ხოლო ჰიდრაქსილის ჯგუფებს შეუძლიათ ურთიერთქმედება სხვა... უჯრედი, სფინგომიელინები არის მიელინის ძირითადი კომპონენტები და ტვინის უჯრედებისა და ნერვული ქსოვილის გარსები. ზოგიერთი პათოლოგიური...სტეროიდები
ქოლესტერინი შეიძლება იყოს ესტერიფიცირებული ჰიდროქსილის ჯგუფში ცხიმოვანი მჟავებით, ქოლესტერინის ეთერების წარმოქმნით. არაესტერიფიცირებული ფორმით, ქოლესტერინი სხვადასხვა უჯრედის მემბრანის ნაწილია.…ვიტამინები
მე-19 საუკუნის შუა ხანებში იდეები კვებითი ღირებულებაცილები, ცხიმები, ნახშირწყლები, მინერალებიდა წყალი. თუმცა ექსპერიმენტული... 1880 წელს ნ.ი. ლუნინმა ჩაატარა კვლევა თაგვებთან, თაგვების ერთ ჯგუფთან... ექიმებმა იაპონიასა და ინდონეზიაში თავიანთ პრაქტიკაში აღნიშნეს, რომ პაციენტები, რომლებიც ძირითადად გაპრიალებულ ბრინჯს უფრო ხშირად მოიხმარდნენ...ზრდასრულთა დიეტა
ამჟამინდელი გამოყენება მნიშვნელოვან როლს ასრულებს ვიტამინის დეფიციტის განვითარებაში. საკვები პროდუქტები, ექვემდებარება... ამრიგად, დიეტა თანამედროვე ადამიანისაკმარისი კალორიული შემცველობით, არა...ცხიმში ხსნადი ვიტამინები
ვიტამინი A ქიმიური სახელია რეტინოლი, კლინიკური სახელია ანტიქსეროფთალმური. რეტინოლი შედგება ბეტა-იონონის რგოლისა და გვერდითი ჯაჭვისგან... საკვებში A ვიტამინი გვხვდება რეტინილ პალმიტატის ეთერების სახით,... A ვიტამინის ბიოლოგიური ეფექტი.წყალში ხსნადი ვიტამინები
ვიტამინი C. ამ ვიტამინის კლინიკური სახელწოდება არის ანტისკორბუტური, ხოლო ქიმიური სახელია...ვიტამინის მსგავსი ნივთიერებები.
PABA (PARAMINOBENZOIC K-TA) 1. მონაწილეობს ფოლიუმის მჟავის წარმოქმნაში, 2. მონაწილეობს მთელი რიგი ფერმენტების წარმოქმნაში,ფერმენტები და არაორგანული კატალიზატორები
ფერმენტები ბიოლოგიური კატალიზატორებია, ძირითადად ცილოვანი ხასიათისა. ფერმენტების როლი ორგანიზმში უზარმაზარია. სხეულის თითოეული უჯრედი შეიცავს მდე... ფერმენტების და არაბიოლოგიური კატალიზატორების ზოგადი მახასიათებლები.ფერმენტების სტრუქტურა
მარტივი ფერმენტები წარმოდგენილია მხოლოდ ცილოვანი ნაწილით (შედგება ამინომჟავებისგან) - პეპსინი, ტრიპსინი, ფოსფატაზები. რთული ფერმენტები წარმოდგენილია: 1. ცილოვანი ნაწილი (შედგება ამინომჟავებისგან) - აპოენზიმი;ბრინჯი. ფერმენტის აქტიური ადგილი
კოენზიმები
კოენზიმები რთული ფერმენტების არაცილოვანი ნაწილია. ისინი იყოფა ორ ჯგუფად: 1. ვიტამინები. 2.არავიტამინური.ფერმენტების თვისებები
1.მაღალი კატალიზური აქტივობა. 2.ფერმენტები, როგორც პროტეინები, ავლენენ თერმოლაბილურ თვისებებს -... ყოველ 10 გრადუს ცელსიუსზე ტემპერატურის მატებასთან ერთად ფერმენტული რეაქციების სიჩქარე იზრდება 1,5 - 2-ჯერ...ფერმენტის მოქმედების სპეციფიკა.
1. აბსოლუტური სპეციფიკა. მას ფლობენ ფერმენტები, რომლებიც მოქმედებენ მხოლოდ ერთ სუბსტრატზე და არ მოქმედებენ სხვა სუბსტრატებზე. ურეაზა აკატალიზებს შარდოვანას ჰიდროლიზს.ფერმენტების ნომენკლატურა
1. ტრივიალური ნომენკლატურა. მაგალითი: პეპსინი, ტრიპსინი. 2. სამუშაო ნომენკლატურა: სახელწოდება S + ტრანსფორმაციის ტიპი + დაბოლოება „აზა“.ფერმენტების კლასიფიკაცია
ფერმენტების კლასიფიკაცია
კატალიზებული რეაქციის ტიპზე დაყრდნობით
ფერმენტების კლასები
ოქსიდორედუქტაზები
ტრანსფერაზები
ჰიდროლაზები
ლიაზეები
იზომერაზები
სინთეტაზები
ფერმენტების მოქმედების მექანიზმი
1 სტადიას ახასიათებს სუბსტრატის დიფუზია ფერმენტში და მათი სტერული ურთიერთქმედება ფერმენტ-სუბსტრატის კომპლექსის წარმოქმნით. ეს ეტაპი... E + S > ESმეორე ეტაპზე ხდება ტრანსფორმაცია E-S კომპლექსიერთ ან რამდენიმე გააქტიურებულ კომპლექსში.
E + S > ES > ES*> ES** > EP
ეს ეტაპი დროში ყველაზე გრძელია. ამ შემთხვევაში სუბსტრატის მოლეკულაში ბმები იშლება და წარმოიქმნება ახალი ბმები, ე.ი. წარმოიქმნება რეაქციის პროდუქტები. აქტივაციის ენერგია მნიშვნელოვნად მცირდება.
მესამე ეტაპზე რეაქციის პროდუქტები გამოიყოფა ფერმენტიდან და გამოიყოფა გარემოში.
EP > E + P
ფერმენტების მიერ ქიმიური რეაქციის დაჩქარება ხდება რეაქტიული ნივთიერებების აქტივაციის ენერგიის მნიშვნელოვანი შემცირების გამო. მთელი რიგი მოლეკულური ეფექტები შესაძლებელს ხდის აქტივაციის ენერგიის შემცირებას.
მოლეკულური ეფექტები
კონცენტრაციის ეფექტი.
სუბსტრატის მოლეკულები კონცენტრირებულია ფერმენტის აქტიური ცენტრის რეგიონში.
სიახლოვის და ორიენტაციის ეფექტი.
ეს არის ფერმენტების დამახასიათებელი თვისება, რაც საშუალებას იძლევა დააჩქაროს სუბსტრატის გარდაქმნა და გაზარდოს რეაქციის სიჩქარე 1000 და 10000-ჯერ. ფერმენტის აქტიური ცენტრის საკონტაქტო უბნები აკავშირებს კონკრეტულად სუბსტრატის მოლეკულებს, აახლოებს მათ და უზრუნველყოფს ორმხრივ ორიენტაციას ისე, რომ ეს სასარგებლო იყოს ფერმენტის კატალიზური ჯგუფების მოქმედებისთვის. სუბსტრატის ეს მოწესრიგებული განლაგება იწვევს აქტივაციის ენერგიის შემცირებას.
დაძაბულობის ეფექტი.
სანამ სუბსტრატი მიმაგრდება ფერმენტის აქტიურ ცენტრს, მისი მოლეკულა მოდუნებულ მდგომარეობაშია. შებოჭვის შემდეგ, სუბსტრატის მოლეკულა იჭიმება და იღებს დაძაბულ, დეფორმირებულ კონფიგურაციას. ამავდროულად, იზრდება ატომთაშორისი ბმების სიგრძე, შესაბამისად, მცირდება აქტივაციის ენერგია.
მჟავა-ტუტოვანი კატალიზი.
კოვალენტური კატალიზი.
იგი შეინიშნება ფერმენტებში, რომლებიც ქმნიან კოვალენტურ კავშირებს აქტიური ადგილისა და სუბსტრატის კატალიზურ ჯგუფებს შორის. შედეგად წარმოიქმნება შუალედური ფერმენტ-სუბსტრატის კომპლექსი, რომელიც არასტაბილურია, ადვილად იშლება და რეაქციის პროდუქტები სწრაფად გამოიყოფა.
გამოწვეული შესაბამისობის ეფექტი.
ის ხსნის ფერმენტის მოქმედების სპეციფიკას. ამ საკითხზე 2 თვალსაზრისია:
ა). ფიშერის ჰიპოთეზა.
ბ). კოშლანდის ინდუცირებული კორესპონდენციის თეორია ავსებდა ფიშერის თეორიას. მისი მიხედვით, ფერმენტის მოლეკულა არის არა ხისტი, არამედ მოქნილი სტრუქტურა. შემდეგ... წინა სექცია უმაღლესი დონის სექცია...- ეს არის კატალიზური აქტივობის დაქვეითება გარკვეული ნივთიერებების - ინჰიბიტორების არსებობისას.

მათი მოქმედების ბუნებიდან გამომდინარე, ინჰიბიტორები იყოფა 2 დიდ ჯგუფად:
შექცევადია ნაერთები, რომლებიც არაკოვალენტურად ურთიერთქმედებენ ფერმენტთან, რითაც წარმოქმნიან კომპლექსს, რომელსაც შეუძლია დისოციაცია.
შეუქცევადია ნაერთები, რომლებსაც შეუძლიათ კონკრეტულად დააკავშირონ ფერმენტის აქტიური ცენტრის გარკვეული ფუნქციური ჯგუფები. ისინი ქმნიან მასთან ძლიერ კოვალენტურ კავშირებს, ამიტომ ასეთი კომპლექსის განადგურება რთულია.
ინჰიბირების სახეები.
მოქმედების მექანიზმის მიხედვით არსებობს შემდეგი ტიპებიინჰიბირება:
კონკურენტული ინჰიბიცია არის ფერმენტული რეაქციის ინჰიბიცია, რომელიც გამოწვეულია ინჰიბიტორის ფერმენტის აქტიურ ცენტრთან შეკავშირებით, რომელიც თავისი სტრუქტურით ახლოსაა სუბსტრატის სტრუქტურასთან. ამ შემთხვევაში, სუბსტრატსაც და ინჰიბიტორსაც შეუძლიათ ფერმენტთან ურთიერთქმედება, მაგრამ ისინი კონკურენციას გაუწევენ ფერმენტის აქტიურ ცენტრს და ნივთიერება, რომელიც უფრო უხვია, შებოჭავს.
მაგალითი:

სუქცინატის დეჰიდროგენაზას რეაქცია.
ამ რეაქციის კონკურენტული ინჰიბიტორია მალონის მჟავა, ამიტომ ორივე მჟავა აკავშირებს ფერმენტის აქტიურ ცენტრს, ხსნარში მათი თანაფარდობის მიხედვით. კონკურენტული ინჰიბიტორის ეფექტის ნაწილობრივ ან მთლიანად მოსაშორებლად საჭიროა სუბსტრატის კონცენტრაციის გაზრდა. ამ შემთხვევაში, მთელი ფერმენტი იქნება ფერმენტ-სუბსტრატის კომპლექსის სახით, ხოლო ფერმენტ-ინჰიბიტორის კომპლექსის პროპორცია მკვეთრად შემცირდება, ამიტომ ფერმენტული რეაქციის სიჩქარე შეიძლება იყოს მაქსიმალური ინჰიბიტორის არსებობის შემთხვევაშიც კი.
ბევრი მედიკამენტებიმოქმედებს როგორც კონკურენტული ინჰიბიტორი. ამავდროულად, ისინი აფერხებენ ბაქტერიული უჯრედების ფუნქციონირებისთვის საჭირო მთელი რიგი ფერმენტების აქტივობას. ამის მაგალითია სულფონამიდების გამოყენება. სხვადასხვა დროს ინფექციური დაავადებებირომლებიც გამოწვეულია ბაქტერიებით, გამოიყენება სულფონამიდური პრეპარატები.
ეს პრეპარატები სტრუქტურულად მსგავსია პარაამინობენზოის მჟავას, რომელსაც ბაქტერიები იყენებენ ფოლიუმის მჟავას სინთეზისთვის, რომელიც აუცილებელია ბაქტერიების ზრდისა და რეპროდუქციისთვის.
სულფონამიდების შეყვანა იწვევს ბაქტერიული ფერმენტების ინჰიბირებას, რომლებიც ასინთეზირებენ ფოლიუმის მჟავას. ამ მჟავის სინთეზის დარღვევა იწვევს მიკროორგანიზმების ზრდის დარღვევას და მათ სიკვდილს.
მთელი ჯგუფი მუშაობს კონკურენტული ინჰიბიტორების პრინციპით სხვადასხვა ნარკოტიკებიარის ანტიქოლინესთერაზები. ისინი წარმოადგენენ ფერმენტ ქოლინესტერაზას კონკურენტულ ინჰიბიტორებს, რომელიც ახორციელებს აცეტილქოლინის ჰიდროლიზს. აცეტილქოლინი უზრუნველყოფს გამტარობას ნერვული იმპულსი. ანტიქოლინესთერაზები კონკურენციას უწევენ აცეტილქოლინს ქოლინესტერაზას ფერმენტის აქტიური ადგილისთვის. შედეგად, აცეტილქოლინის დაშლა თრგუნავს, ის გროვდება ორგანიზმში, რაც იწვევს ნერვული იმპულსების გამტარობის დარღვევას.
არაკონკურენტული ინჰიბიცია არის ფერმენტული რეაქციის დათრგუნვა, რომელიც გამოწვეულია ინჰიბიტორის ზემოქმედებით სუბსტრატის კატალიზურ გარდაქმნაზე. ამ შემთხვევაში ინჰიბიტორი არ ახდენს გავლენას ფერმენტის სუბსტრატთან შეკავშირებაზე. არაკონკურენტულ ინჰიბიტორს შეუძლია დაუკავშირდეს ფერმენტის აქტიური უბნის კატალიზურ ჯგუფებს ან ფერმენტის აქტიური უბნის გარეთ, მაგრამ ამით ის ცვლის ფერმენტის კონფორმაციას და გავლენას ახდენს მისი აქტიური ადგილის კატალიზურ ადგილზე. არაკონკურენტული ინჰიბიციით შესაძლებელია სამიანი, არააქტიური კომპლექსის წარმოქმნა.

არაკონკურენტული დათრგუნვის სქემა
ციანიდი მოქმედებს როგორც არაკონკურენტული ინჰიბიტორი. ისინი ძლიერად უკავშირდებიან რკინის იონებს, რომლებიც კატალიზური ჰემის ფერმენტის, ციტოქრომ ოქსიდაზას ნაწილია. ეს ფერმენტი არის რესპირატორული ჯაჭვის ერთ-ერთი კომპონენტი. სასუნთქი ჯაჭვის დაბლოკვა აჩერებს მის მუშაობას, რაც იწვევს ორგანიზმში მყისიერ სიკვდილს.
არაკონკურენტული ინჰიბიტორის მაგალითია მძიმე ლითონის მარილების მოქმედება. ისინი ბლოკავენ -SH ჯგუფებს, რომლებიც ფერმენტის კატალიზური ადგილის ნაწილია. ამ შემთხვევაში წარმოიქმნება ფერმენტ-ინჰიბიტორის კომპლექსი. მას შეუძლია სუბსტრატის მიმაგრება, მაგრამ სუბსტრატის შემდგომი ტრანსფორმაცია არ ხდება, რადგან ფერმენტის კატალიზური ჯგუფები დაბლოკილია. რეაქცია არაპროდუქტიულია. ძალიან რთულია არაკონკურენტული ინჰიბიტორის ეფექტის მოხსნა, რადგან ლითონის იონები ძალიან მჭიდროდ უკავშირდება ფერმენტის აქტიურ ადგილს. ამ ინჰიბიტორის ეფექტი შეიძლება შეიცვალოს მხოლოდ სპეციალური ნივთიერებები- რეაქტივატორები.
3. სუბსტრატის ინჰიბირება არის ფერმენტული რეაქციის დათრგუნვა, რომელიც გამოწვეულია სუბსტრატის სიჭარბით. ამ შემთხვევაში წარმოიქმნება ფერმენტ-სუბსტრატის კომპლექსი, რომელიც არ განიცდის კატალიზურ გარდაქმნებს, რადგან ფერმენტის მოლეკულას არააქტიურს ხდის. სუბსტრატის ინჰიბიტორის ეფექტი იხსნება სუბსტრატის კონცენტრაციის შემცირებით.
4. ალოსტერიული დათრგუნვა დამახასიათებელია მეოთხეული სტრუქტურის მქონე ფერმენტებისთვის, რომელთა მოლეკულა შედგება რამდენიმე ერთეულისაგან (პროტომერები). ალოსტერულ ფერმენტებს შეიძლება ჰქონდეთ 2 ან მეტი ერთეული. ამ შემთხვევაში, ერთს აქვს კატალიზური ცენტრი და ეწოდება კატალიზური, ხოლო მეორეს აქვს ალოსტერიული ცენტრი და ეწოდება მარეგულირებელი. ალოსტერიული ინჰიბიტორის არარსებობის შემთხვევაში, სუბსტრატი აკავშირებს კატალიზურ ადგილს და ნორმალური კატალიზური რეაქცია მიმდინარეობს. როდესაც ჩნდება ალოსტერიული ინჰიბიტორი, ის ერთვის მარეგულირებელ ერთეულს, ე.ი. ალოსტერიულ ცენტრამდე და ცვლის ფერმენტის ცენტრის კონფორმაციას, რის შედეგადაც ფერმენტის აქტივობა მცირდება.
ნახშირწყლების მეტაბოლიზმი
1. ცხოველის ორგანიზმის ძირითადი ნახშირწყლები, მათი ბიოლოგიური როლი. 2. საჭმლის მომნელებელი სისტემის ორგანოებში ნახშირწყლების გარდაქმნა. 3. ქსოვილებში გლიკოგენის ბიოსინთეზი და დაშლა.ნახშირწყლების ბიოლოგიური როლი
1. ენერგია. როდესაც 1 გ ნახშირწყლები იჟანგება საბოლოო პროდუქტებამდე (CO2 და H2O), გამოყოფს... 2. სტრუქტურული.საჭმლის მომნელებელ ტრაქტში ნახშირწყლების გარდაქმნა
საჭმლის მომნელებელი ტრაქტი ადამიანის ორგანიზმისთვის საკვების ძირითადი ნახშირწყლებია: სახამებელი, გლიკოგენი,... საკვებიდან მიღებული სახამებელი (გლიკოგენი) პირის ღრუსგანიცდის ჰიდროლიზს სანერწყვე ალფა-ამილაზას მიერ,...გლიკოგენის ბიოსინთეზი და დაშლა
გლიკოგენური დაავადებები. აღმოჩნდა, რომ გლიკოგენის სინთეზირება შესაძლებელია თითქმის ყველა...ანაერობული გლიკოლიზი
1. ანაერობულ პირობებში 2. აერობულ პირობებში. ანაერობული გლიკოლიზი (გლიკოგენოლიზი) ხდება უჯრედების ციტოპლაზმაში. გლუკოზის ან გლიკოგენის გლუკოზის ნარჩენების დაჟანგვა...ბრინჯი. გლუკოზის აერობული დაშლა
აერობული გლიკოლიზი (ჰექსოზა დიფოსფატის გზა)
ეს არის ქსოვილებში ნახშირწყლების აერობული კატაბოლიზმის კლასიკური გზა, გრძელდება ციტოპლაზმაში პირუვატის წარმოქმნის სტადიამდე და მთავრდება მიტოქონდრიაში... როდესაც ჟანგბადი იწყებს უჯრედებში შეღწევას, ხდება ანაერობული დათრგუნვა... 1. გლიცერალდეჰიდი- 3-ფოსფატიბრინჯი. გლუკოზის აერობული დაშლა
ჰექსოზა მონოფოსფატის გზა
ქსოვილებში, რეაქციების ქიმია. გლუკოზის დაჟანგვა ამ გზის გასწვრივ ხდება უჯრედების ციტოპლაზმაში და წარმოდგენილია... გლუკოზის დაჟანგვის ამ გზის ბიოლოგიური როლი პირველ რიგში დაკავშირებულია ორი ნივთიერების გამომუშავებასთან:გლუკონეოგენეზი
ადამიანის ორგანიზმისთვის გლუკოზის ძირითადი წყაროა: 1. დიეტური ნახშირწყლები; 2. ქსოვილის გლიკოგენი;ადამიანის სხეულის ძირითადი ლიპიდები და მათი ბიოლოგიური როლი.
ლიპიდები ძირითადი საკვებია. ისინი ადამიანის ორგანიზმში შედიან მცენარეული და ცხოველური წარმოშობის პროდუქტებით. ყოველდღიურად... ადამიანის ორგანიზმში ლიპიდები წარმოდგენილია: 1. სტრუქტურული ლიპიდებით (ქოლესტერინი, ფოსფოლიპიდები, გლიკოლიპიდები).ლიპიდების მონელება, ცხიმის რესინთეზი
პირის ღრუში საკვებიდან მომდინარე ლიპიდები ექვემდებარება მხოლოდ მექანიკურ დამუშავებას. LIPOLYTIC ფერმენტები პირის ღრუში არ წარმოიქმნება... 1. ნაღვლის მჟავების არსებობა. 2. ფერმენტების არსებობა.სისხლის ლიპოპროტეინები
მათი სტრუქტურის მიხედვით, ლიპოპროტეინის მიცელებს აქვთ გარე შრე და ბირთვი. გარე შრე წარმოიქმნება ცილების, ფოსფოლიპიდების და ქოლესტერინისგან, რომლებსაც აქვთ... სისხლში არის ლიპოპროტეინების 4 კლასი, რომლებიც ერთმანეთისგან განსხვავდება... 1. ქილომიკრონებით. ისინი წარმოიქმნება ნაწლავის კედელში და აქვთ ყველაზე დიდი ნაწილაკების ზომა.უმაღლესი ცხიმოვანი მჟავების დაჟანგვა
ცხიმოვანი ქსოვილის უჯრედებში TAG რღვევა ხდება ლიპაზების მონაწილეობით. ლიპაზა არის არააქტიური ფორმით, მას ააქტიურებენ ჰორმონები (ადრენალინი,... IVF ალბუმინების დახმარებით სისხლით ტრანსპორტირდება ქსოვილების, ორგანოების უჯრედებში, სადაც... უმაღლესი ცხიმოვანი მჟავების დაჟანგვა.გლიცეროლის დაჟანგვა
გლიცეროლის დაჟანგვა ქსოვილებში მჭიდრო კავშირშია გლიკოლიზთან, რომელიც მოიცავს გლიცეროლის ცვლის მეტაბოლიტებს შემდეგი სქემის მიხედვით:

გლიცეროლის დაჟანგვის შედეგად წარმოიქმნება შემდეგი საბოლოო პროდუქტები:
CO2 კონვერტაციის ეტაპზე:
პირუვატი
იზოციტრატი
ალფა-კეტოგლუტარატი
H 2 O ტრანსფორმაციის ეტაპზე:
ალფა-გლიცეროფოსფატი
გლიცერალდეჰიდი-3-ფოსფატი
ფოსფოგლიცერატი
პირუვატი
იზოციტრატი
ალფა-კეტოგლუტარატი
სუქცინატი
მალატა
ATP გამოუშვეს რეაქციების გამო
ა) სუბსტრატის ფოსფორილირება ტრანსფორმაციის ეტაპებზე:
დიფოსფოგლიცერატი
ფოსფენოლპირუვატი
სუქცინილ-CoA
ბ) ოქსიდაციური ფოსფორილირება ტრანსფორმაციის ეტაპებზე:
ალფა-გლიცერაფოსფატი
გლიცერალდეჰიდი-3 ფოსფატი
პირუვატი
იზოციტრატი
ალფა-კეტოგლუტარატი
სუქცინატი
მალათა
ერთი გლიცეროლის მოლეკულის დაჟანგვის მთლიანი ენერგეტიკული ეფექტი უდრის 22 ATP.
IVH ბიოსინთეზი ქსოვილებში
VFA-ს ბიოსინთეზის პირობებია: 1. ACETYL-CoA, ATP, CO2, H2O, NADP*H2, 2. სპეციალური გადამზიდავი ცილების (HS-ACP) არსებობა. 3.სინთეზის ფერმენტების არსებობა.ქოლესტერინის გაცვლა
ქოლესტერინი არის სტეროიდების სინთეზის წინამორბედი: ნაღვლის მჟავები, სტეროიდული ჰორმონები, ვიტამინი D3. ქოლესტერინი არის აუცილებელი…ცილების მონელება
საკვების ცილები პროტეოლიზური ფერმენტების (კლასი - ჰიდროლაზები, ქვეკლასი - პეპტიდაზები) მოქმედებით განიცდიან ჰიდროლიზურ დაშლას. ამ ფერმენტების უმეტესობა იწარმოება არააქტიური ფორმით, ე.ი. სახით... პროენზიმები წარმოიქმნება კუჭის ან ნაწლავების ლორწოვანი გარსის, პანკრეასის უჯრედებში და...ამინომჟავების გაფუჭება, დამპალი პროდუქტების განეიტრალება
ამინომჟავები, რომლებიც არ შეიწოვება, შედიან მსხვილ ნაწლავში, სადაც განიცდიან დაშლას. დეკარბოქსილაციური რეაქციები: ამინომჟავები დაშლის პროცესია...ამინომჟავების მეტაბოლიზმი
უჯრედში ამინომჟავების წყაროა: 1. საკვების ცილები საჭმლის მომნელებელ ორგანოებში მათი ჰიდროლიზის შემდეგ; 2. სინთეზი არაარსებითი ამინომჟავები;ბრინჯი. გლუტამინის მჟავას ოქსიდაციური დეამინირება
არაპირდაპირი დემინაცია
დარჩენილი ამინომჟავები გადიან ამ ტიპის დეამინაციას, მაგრამ ტრანსამინაციის სტადიის გავლით ალფა-კეტოგლუტარის მჟავით. გლუტამინის მჟავა (ამ რეაქციის პროდუქტი) შემდეგ განიცდის ჟანგვითი დეამინაციას.
ამიაკის განეიტრალების გზები
ამიაკი წარმოიქმნება სხვადასხვა ქსოვილებში. მისი კონცენტრაცია სისხლში უმნიშვნელოა, რადგან ეს არის ტოქსიკური ნივთიერება (0,4 - 0,7 მგ/ლ). განსაკუთრებით... ამიაკის განადგურების გზები.ბრინჯი. გლუტამინის ფორმირება
გლუტამინი და ასპარაგინი არატოქსიკური ნივთიერებებია. მათ სხეულში ამიაკის სატრანსპორტო ფორმას უწოდებენ. ისინი არ აღწევენ გარსებში და თირკმელებში იშლება ამინომჟავებად და ამიაკად.
2. ალფა-კეტოგლუტარის მჟავის რედუქციური ამინება

3. ამონიუმის მარილების წარმოქმნა
4. შარდოვანას სინთეზი არის ამიაკის ნეიტრალიზაციის მთავარი გზა - ORNITHINE CYCLE.
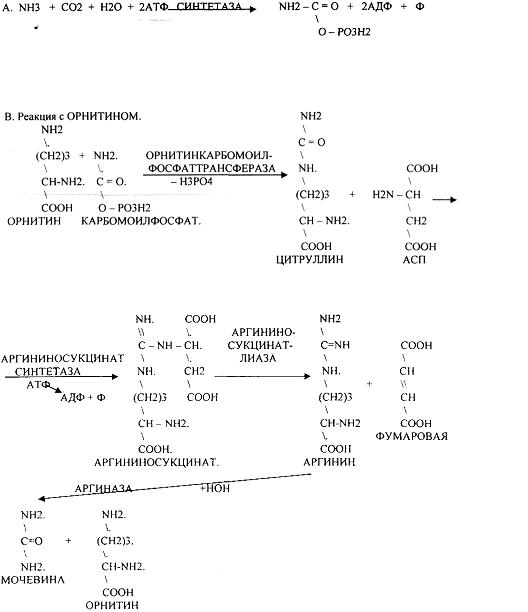
არგინაზას აქვს აბსოლუტური სპეციფიკა და გვხვდება მხოლოდ ღვიძლში. შარდოვანა შეიცავს აზოტის ორ ატომს: ერთი მოდის ამიაკიდან, მეორე კი ASP-დან.
შარდოვანა წარმოიქმნება მხოლოდ ღვიძლში.
ციკლის პირველი ორი რეაქცია (ციტრულინისა და არგინოსუკცინატის წარმოქმნა) ხდება მიტოქონდრიაში, დანარჩენი კი ციტოპლაზმაში.
ორგანიზმი გამოიმუშავებს 25 გ შარდოვანას დღეში. ეს მაჩვენებელი ახასიათებს ღვიძლის შარდოვანას ფორმირების ფუნქციას. შარდოვანა ღვიძლიდან ხვდება თირკმელებში, სადაც გამოიყოფა ორგანიზმიდან როგორც საბოლოო პროდუქტიაზოტის მეტაბოლიზმი.
- 6.1. სასიგნალო მოლეკულები
- 6.2. ჰიპოთალამუსის ჰორმონები
- 6.3. ჰიპოფიზის ჰორმონები
- 6.4. ფარისებრი ჯირკვლის ჰორმონები
- 6.5. პარათირეოიდული ჯირკვლების ჰორმონები
- 6.6. გონადალური ჰორმონები
- 6.7. თირკმელზედა ჯირკვლის ჰორმონები
- 6.8. პანკრეასის ჰორმონები
მეტაბოლიზმის რეგულირება
მეტაბოლიზმის და უჯრედული ფუნქციების რეგულირების ძირითადი ამოცანები: 1. უჯრედშიდა და უჯრედშორისი კოორდინაცია. მეტაბოლური პროცესები; 2. „უსაქმური“ მეტაბოლური ციკლების გამორიცხვა, რომელთა პროდუქტებზე მოთხოვნა არ არის;სასიგნალო მოლეკულები
ჰიპოთალამუსის ჰორმონები
ჰიპოთალამუსი არის ლიმბური სისტემის კომპონენტი და ერთგვარი "გამომავალი არხი". ეს არის დიენცეფალონის მონაკვეთი, რომელიც აკონტროლებს სხვადასხვა პარამეტრებს... ერთი მხრივ, ის დაკავშირებულია ცენტრალურთან. ნერვული სისტემამეორეს მხრივ, ჰიპოფიზის ჯირკვალთან ერთად ნეირონების აქსონებისა და სისტემის...ჰიპოფიზის ჰორმონები
ჰიპოფიზის ჰორმონები
ჰიპოფიზის ჯირკვალი იყოფა წინა (ადენოჰიპოფიზი) და უკანა წილები (ნეიროჰიპოფიზი).ფარისებრი ჯირკვლის ჰორმონები
ფარისებრი ჯირკვლის ჰორმონები
ფარისებრი ჯირკვალი ასინთეზირებს ჰორმონებს, რომლებიც წარმოადგენენ ტიროზინის იოდირებულ წარმოებულებს (იოდოთირონინს). მათ შორისაა ტრიიოდთირონინი (3, 5, 3? - ტრიიოდთირონინი, T 3) და თიროქსინი (3, 5, 3?, 5?, - ტეტრაიოდთირონინი, T 4).
ბრინჯი. იოდოთირონინები
იოდთირონინის ბიოსინთეზი
ტიროზინის იოდირება და იოდთირონინის წარმოქმნა ხდება რამდენიმე ეტაპად: 1. იოდის ტრანსპორტირება უჯრედებში. ფარისებრი ჯირკვალი; 2. იოდის დაჟანგვა;ეფექტი მეტაბოლურ პროცესებზე
ცილის მეტაბოლიზმი
ფიზიოლოგიურ კონცენტრაციებში იოდოთირონინი აძლიერებს ცილის ბიოსინთეზს, მაგრამ მაღალი კონცენტრაციავლინდება მათი კატაბოლური მოქმედება ცილების სინთეზზე.
ნახშირწყლების მეტაბოლიზმი
ღვიძლში, ამ ჰორმონების გავლენის ქვეშ, იზრდება გლუკოზის დაშლის და გლიკოგენის მობილიზაციის სიჩქარე.
ლიპიდური მეტაბოლიზმი
ფარისებრი ჯირკვლის ჰორმონების ზემოქმედებით მატულობს Na+, K+ - ატფ-აზას აქტივობა, რაც იწვევს უჯრედში ატფ-ის დაქვეითებას და, შესაბამისად,... ნორმალურ კონცენტრაციებში იოდოთირონინი ასტიმულირებს ზრდის პროცესებს და უჯრედულს. ..ფარისებრი ჯირკვლის ჰორმონის სეკრეციის დარღვევა
ჰიპოსეკრეცია
მოზრდილებში ფარისებრი ჯირკვლის ჰორმონების ნაკლებობის მძიმე გამოვლინებაა მიქსედემა. ამ შემთხვევაში რღვევა ნელდება... ჰიპოთირეოზი შეიძლება მოხდეს იოდის არასაკმარისი მიღების გამოც...ჰიპერსეკრეცია
პარათირეოიდული ჯირკვლების ჰორმონები
პარათირეოიდული ჯირკვლების ჰორმონები
PTH რეცეპტორები განლაგებულია თირკმლის უჯრედებისა და ძვლოვანი ქსოვილის ზედაპირზე (ოსტეობლასტები, ოსტეოციტები). მოქმედების მექანიზმი დამოკიდებულია cAMP-ზე. ჰორმონის მოქმედება მიზნად ისახავს კალციუმის იონების კონცენტრაციის გაზრდას და სისხლში ფოსფატების კონცენტრაციის შემცირებას.რას ვიზამთ მიღებულ მასალასთან:
თუ ეს მასალა თქვენთვის სასარგებლო იყო, შეგიძლიათ შეინახოთ იგი თქვენს გვერდზე სოციალურ ქსელებში:





