পেশী এবং পেশী সংকোচনের বায়োকেমিস্ট্রি। পেশীর জৈব রসায়ন সংক্ষেপে পেশী সংকোচনের বায়োকেমিস্ট্রি
এই প্রক্রিয়ায় একটি গুরুত্বপূর্ণ ভূমিকা ক্যালসিয়াম আয়ন এবং সারকোপ্লাজমিক প্রোটিন দ্বারা অভিনয় করা হয় - ক্যালসকোয়েস্ট্রিন এবং ক্যালসিয়ামের জন্য একটি উচ্চ সম্বন্ধযুক্ত প্রোটিন। সারকোপ্লাজমিক রেটিকুলামের ঝিল্লি পেশী ফিলামেন্টকে ঘিরে থাকে। এই প্রোটিনগুলি ভিতরের ঝিল্লিতে এসপিআর-এর সিস্টারনে অবস্থিত, যেখানে Ca 2+ আয়নগুলি আবদ্ধ হয়। Calsequestrin - অ্যাসিড গ্লাইকোপ্রোটিন (MM 45,000 Da), 45 Ca 2+ আয়ন সংযুক্ত করতে সক্ষম, ক্যালসিয়ামের জন্য উচ্চ সম্বন্ধযুক্ত একটি প্রোটিন (MM 55,000 Da) 25 Ca 2+ আয়নকে আবদ্ধ করে। ট্যাঙ্কগুলি থেকে Ca 2+ এর স্থানান্তর সরল প্রসারণের মাধ্যমে ঘনত্ব গ্রেডিয়েন্ট বরাবর ঘটে; সাইটোপ্লাজম থেকে সিস্টারনে Ca 2+ স্থানান্তর Ca 2+-নির্ভর ATPase এবং ATP-এর অংশগ্রহণের সাথে একটি গ্রেডিয়েন্টের বিপরীতে। বিশ্রামে, সক্রিয় পরিবহন ব্যবস্থা সিস্টারনে ক্যালসিয়াম সঞ্চয় করে। পেশী সংকোচন আগমনের সাথে শুরু হয় কর্ম সম্ভাব্যমোটর স্নায়ুর শেষ প্লেটে। অ্যাসিটাইলকোলিন সিন্যাপসে নিঃসৃত হয়, যা পেশী ফাইবারে পোস্টসিনাপটিক রিসেপ্টরগুলির সাথে আবদ্ধ হয়। আরও, অ্যাকশন পটেনশিয়াল টি-সিস্টেমের ট্রান্সভার্স টিউবুলে সারকোলেমা বরাবর প্রচার করে। জেড-লাইনগুলির অঞ্চলে, সংকেতটি ট্রান্সভার্স টিউবুল থেকে সারকোপ্লাজমিক রেটিকুলামের সিস্টারনে প্রেরণ করা হয়।
সিস্টার মেমব্রেনের ডিপোলারাইজেশন ক্যালসিয়ামের মুক্তি এবং পেশী সংকোচনের সূত্রপাত ঘটায়। ক্যালসিয়াম ট্রপোনিনের C সাবইউনিটের সাথে আবদ্ধ হয়। এটি সম্পূর্ণ ট্রপোনিন অণুর গঠন পরিবর্তন করে - সাবুনিট I মায়োসিনের সাথে অ্যাক্টিনের মিথস্ক্রিয়ায় হস্তক্ষেপ করা বন্ধ করে দেয়; টি সাবুনিটের গঠনের পরিবর্তন ট্রপোমায়োসিনে প্রেরণ করা হয়। ট্রোপোমায়োসিন তারপর 20° ঘোরে এবং মায়োসিনের সাথে আবদ্ধ হওয়ার জন্য অ্যাক্টিনে পূর্বে বন্ধ হওয়া কেন্দ্রগুলি খোলে। মায়োসিন হেড, যা বিশ্রামে ADP + P এবং + myosin এর একটি কমপ্লেক্স, অ্যাক্টিনের সাথে লম্বভাবে সংযুক্ত থাকে এবং এই কমপ্লেক্সের (ট্রান্সভার্স ব্রিজ গঠন) জন্য অ্যাক্টিনের একটি উচ্চ সম্পর্ক রয়েছে। অ্যাক্টিন সংযুক্ত করার ফলে মায়োসিন থেকে ADP এবং Pn দ্রুত মুক্তি পায়। এটি গঠনের পরিবর্তনের দিকে নিয়ে যায়, এবং মায়োসিন মাথা 45° (স্ট্রোক) ঘোরে। অ্যাক্টিন-বাউন্ড মাথার ঘূর্ণনের ফলে মায়োসিনের তুলনায় পাতলা ফিলামেন্ট সরে যায়। ATP প্রস্থান করা ADP এবং Fn এর পরিবর্তে মায়োসিন মাথার সাথে পুনরায় সংযুক্ত করা হয়, M + ATP কমপ্লেক্স গঠন করে। অ্যাক্টিন এর জন্য একটি কম সখ্যতা রয়েছে, যা মায়োসিন মাথার বিচ্ছিন্নতা সৃষ্টি করে (ট্রান্সভার্স ব্রিজ ফেটে যাওয়া)। এটি আবার পাতলা থ্রেডে লম্ব হয়ে যায়। মায়োসিনের মাথায়, যা অ্যাক্টিনের সাথে যুক্ত নয়, এটিপি হাইড্রোলাইসিস ঘটে। ADP + P n + myosin কমপ্লেক্স আবার গঠিত হয়, এবং সবকিছু পুনরাবৃত্তি হয়। মায়োসিনে ATP-এর সংযোজন এবং ATP-এর হাইড্রোলাইসিস খুব দ্রুত ঘটে, কিন্তু ADP এবং Pn-এর হাইড্রোলাইসিস পণ্যগুলি ধীরে ধীরে মায়োসিন থেকে বিভক্ত হয়ে যায়।
মোটর ইমপালসের ক্রিয়া শেষ হওয়ার পরে, Ca 2+ Ca 2+-নির্ভর ATPase-এর সাহায্যে সারকোপ্লাজমিক রেটিকুলামে প্রবেশ করে। ট্রপোনিন কমপ্লেক্স থেকে ক্যালসিয়াম ত্যাগ করার ফলে ট্রপোমায়োসিনের স্থানচ্যুতি ঘটে এবং অ্যাক্টিনের সক্রিয় কেন্দ্রগুলি বন্ধ হয়ে যায়, এটি মায়োসিনের সাথে যোগাযোগ করতে অক্ষম করে তোলে - পেশী শিথিল হয়।
পেশী স্বাস্থ্যের জন্য গুরুত্বপূর্ণ নিউরোমাসকুলার সংক্রমণ প্রক্রিয়া। এক)মায়াস্থেনিয়া গ্র্যাভিসের সাথে, তাদের নিজস্ব অ্যাসিটাইলকোলিন রিসেপ্টরগুলির বিরুদ্ধে অ্যান্টিবডিগুলি রক্তে পাওয়া যায়, যা পেশী দুর্বলতার দ্বারা প্রকাশিত হয়।
2).
অনেকগুলি ওষুধ (অ্যাট্রোপিন, সাকসিনাইলকোলিন, কিউরে বিষ) বাধা দেয় রিসেপ্টর প্রোটিন, জএটি নিউরোমাসকুলার সঞ্চালন ব্লক করে।
3).
ওষুধ (নিওস্টিগমাইন, ইজারিন) বাধা দেয় অ্যাসিটাইলকোলিন এস্টারেজযার ফলে এসিটাইলকোলিনের ক্রিয়া বৃদ্ধি পায়।
4).
আরও শক্তিশালী এনজাইম ইনহিবিটার হল জৈব ফ্লুরোফসফেটস। তারা অ্যাসিটাইলকোলিনস্টেরেজের সাথে একটি শক্তিশালী বন্ধন তৈরি করে এবং শ্বাসকষ্টের কারণে মৃত্যু ঘটায়। এগুলো নার্ভ পয়জন- ট্যাবুন, সারিন।
সংকোচনের সময় পেশীতে ঘটতে থাকা চক্রীয় জৈব রাসায়নিক বিক্রিয়াগুলি "মাথা" - পুরু প্রোটোফাইব্রিল এবং প্রোট্রুশনগুলির মায়োসিন অণুর বৃদ্ধি - পাতলা প্রোটোফাইব্রিলের সক্রিয় কেন্দ্রগুলির মধ্যে আনুগত্যের পুনরাবৃত্তি এবং ধ্বংস নিশ্চিত করে। আঠালো গঠন এবং মায়োসিন ফিলামেন্ট বরাবর অ্যাক্টিন ফিলামেন্টের প্রচারের জন্য সুনির্দিষ্ট নিয়ন্ত্রণ এবং উল্লেখযোগ্য শক্তি ব্যয় উভয়ই প্রয়োজন। বাস্তবে, ফাইবার সংকোচনের মুহুর্তে, প্রতিটি সক্রিয় কেন্দ্রে প্রতি মিনিটে প্রায় 300 আনুগত্য তৈরি হয় - একটি লেজ।
যেমনটি আমরা আগে উল্লেখ করেছি, শুধুমাত্র ATP-এর শক্তি সরাসরি পেশী সংকোচনের যান্ত্রিক কাজে রূপান্তরিত হতে পারে। মায়োসিনের এনজাইমেটিক কেন্দ্র দ্বারা হাইড্রোলাইজ করা ATP সম্পূর্ণ প্রোটিন মায়োসিনের সাথে একটি জটিল গঠন করে। এটিপি-মায়োসিন কমপ্লেক্সে, শক্তিতে পরিপূর্ণ, মায়োসিন তার গঠন পরিবর্তন করে এবং এর সাথে বাহ্যিক "মাত্রা" এবং এইভাবে, মায়োসিন ফিলামেন্টের বৃদ্ধিকে ছোট করার জন্য যান্ত্রিক কাজ করে।
একটি বিশ্রাম পেশীতে, মায়োসিন এখনও এটিপির সাথে যুক্ত থাকে, তবে এটিপির হাইড্রোলাইটিক ক্লিভেজ ছাড়াই Mg ++ আয়নের মাধ্যমে। বিশ্রামে মায়োসিন এবং অ্যাক্টিনের মধ্যে আনুগত্যের গঠন ট্রপোনিনের সাথে ট্রপোমায়োসিনের জটিল দ্বারা প্রতিরোধ করা হয়, যা অ্যাক্টিনের সক্রিয় কেন্দ্রগুলিকে অবরুদ্ধ করে। অবরোধ বজায় রাখা হয় এবং Ca ++ আয়ন আবদ্ধ থাকা অবস্থায় ATP বিভক্ত হয় না। যখন একটি স্নায়ু আবেগ একটি পেশী ফাইবারে আসে, তখন এটি মুক্তি পায় পালস ট্রান্সমিটার- নিউরোহরমোন অ্যাসিটাইলকোলিন Na + আয়নগুলির সাথে, সারকোলেমার অভ্যন্তরীণ পৃষ্ঠের নেতিবাচক চার্জ নিরপেক্ষ হয় এবং এর বিধ্বংসীকরণ ঘটে। এই ক্ষেত্রে, Ca ++ আয়ন নির্গত হয় এবং ট্রোপোনিনের সাথে আবদ্ধ হয়। পরিবর্তে, ট্রপোনিন তার চার্জ হারায়, যে কারণে সক্রিয় কেন্দ্রগুলি - অ্যাক্টিন ফিলামেন্টের প্রোট্রুশনগুলি মুক্তি পায় এবং অ্যাক্টিন এবং মায়োসিনের মধ্যে আনুগত্য দেখা দেয় (যেহেতু পাতলা এবং পুরু প্রোটোফাইব্রিলের ইলেক্ট্রোস্ট্যাটিক বিকর্ষণ ইতিমধ্যে সরানো হয়েছে)। এখন, Ca ++ এর উপস্থিতিতে ATP মায়োসিনের এনজাইমেটিক কার্যকলাপের কেন্দ্রের সাথে মিথস্ক্রিয়া করে এবং বিভক্ত হয় এবং রূপান্তরিত কমপ্লেক্সের শক্তি আঠালো কমাতে ব্যবহৃত হয়। উপরে বর্ণিত আণবিক ঘটনাগুলির চেইনটি একটি বৈদ্যুতিক প্রবাহের অনুরূপ যা একটি মাইক্রোক্যাপাসিটর রিচার্জ করে, এর বৈদ্যুতিক শক্তি অবিলম্বে ঘটনাস্থলে যান্ত্রিক কাজে রূপান্তরিত হয় এবং আপনাকে আবার রিচার্জ করতে হবে (যদি আপনি এগিয়ে যেতে চান)।
আনুগত্য ফেটে যাওয়ার পরে, এটিপি বিভক্ত হয় না, তবে আবার মায়োসিনের সাথে একটি এনজাইম-সাবস্ট্রেট কমপ্লেক্স গঠন করে:
M–A + ATP -----> M – ATP + Aবা
M-ADP-A + ATP ----> M-ATP + A + ADP
যদি এই মুহুর্তে একটি নতুন স্নায়ু আবেগ আসে, তবে "রিচার্জিং" প্রতিক্রিয়াগুলি পুনরাবৃত্তি হয়, যদি পরবর্তী আবেগ না আসে, পেশী শিথিল হয়। শিথিলকরণের সময় সংকুচিত পেশীটি তার আসল অবস্থায় ফিরে আসা পেশী স্ট্রোমার প্রোটিনের স্থিতিস্থাপক শক্তি দ্বারা সরবরাহ করা হয়। পেশী সংকোচনের আধুনিক অনুমানকে সামনে রেখে, বিজ্ঞানীরা পরামর্শ দেন যে সংকোচনের মুহুর্তে, অ্যাক্টিন ফিলামেন্টগুলি মায়োসিন ফিলামেন্টগুলির সাথে স্লাইড করে এবং সংকোচনযোগ্য প্রোটিনের স্থানিক কাঠামোর পরিবর্তনের কারণে (হেলিক্সের আকারে পরিবর্তন) তাদের সংক্ষিপ্তকরণও সম্ভব।
বিশ্রামে, এটিপির একটি প্লাস্টিকাইজিং প্রভাব রয়েছে: মায়োসিনের সাথে একত্রিত হয়ে, এটি অ্যাক্টিনের সাথে এর আনুগত্য গঠনে বাধা দেয়। পেশী সংকোচনের সময় বিভাজন, এটিপি আনুগত্য সংক্ষিপ্তকরণের প্রক্রিয়ার জন্য শক্তি সরবরাহ করে, সেইসাথে "ক্যালসিয়াম পাম্প" - Ca ++ আয়ন সরবরাহের কাজ। পেশীতে এটিপির বিভাজন খুব উচ্চ গতিতে ঘটে: আপ প্রতি মিনিটে 1 গ্রাম পেশী প্রতি 10 মাইক্রোমোল। যেহেতু পেশীতে ATP-এর মোট মজুদ ছোট (সেগুলি সর্বাধিক শক্তির সাথে শুধুমাত্র 0.5-1 সেকেন্ডের কাজের জন্য যথেষ্ট হতে পারে), স্বাভাবিক পেশী কার্যকলাপ নিশ্চিত করার জন্য, এটিপি বিভক্ত হওয়ার মতো একই হারে পুনরুদ্ধার করতে হবে।
বক্তৃতা নং 4. পেশী সংকোচনের জন্য শক্তি, পেশী কাজের সময় ঘটে বায়োকেমিক্যাল প্রক্রিয়া।
রেসকিউ রিসিন্থেসিস।
বিশেষত, শুধুমাত্র ATP রাসায়নিক শক্তি (এর মুক্ত অংশ, যা ফসফেট বন্ধনে থাকে) যান্ত্রিক শক্তিতে (উড়ান, চলমান এবং স্লাইডিং) রূপান্তর করতে পারে। সে শক্তি প্রদান করেআনুগত্য সংক্ষিপ্তকরণ প্রক্রিয়া, যথাক্রমে, সামগ্রিক পেশী সংকোচনএবং সংকোচনের সাথে জড়িত Ca ++ আয়ন গঠনের জন্য শক্তি সরবরাহ করে)। একটি জীবন্ত কোষ ক্রমাগত প্রায় 0.25% স্তরে ATP-এর একটি কার্যক্ষম ঘনত্ব বজায় রাখে, যার মধ্যে নিবিড় পেশীর কাজ সহ। যদি (বিপাকীয় ব্যাধিগুলির ক্ষেত্রে) এটিপির ঘনত্বের বৃদ্ধি ঘটে, তবে পেশীর সংকোচনশীলতা বিঘ্নিত হবে (এটি একটি "ন্যাকড়া" এর মতো দেখাবে), যদি হ্রাস ঘটে তবে একটি কঠোরতা দেখা দেবে - একটি অবস্থা ক্রমাগত নন-পাসিং সংকোচন ("পেট্রিফিকেশন")। ATP এর কার্যকারী ঘনত্ব এক সেকেন্ড শক্তিশালী কাজের জন্য যথেষ্ট (3-4 একক সংকোচন)। দীর্ঘায়িত পেশী ক্রিয়াকলাপের সময়, এটিপির কার্যকারী ঘনত্ব এটি পুনরুদ্ধার করার প্রতিক্রিয়ার কারণে বজায় রাখা হয়। বিপাক প্রক্রিয়ায় স্বাভাবিক (দীর্ঘমেয়াদী) পেশীর কাজ নিশ্চিত করার জন্য, এটিপি বিভক্ত হওয়ার মতো একই হারে পুনরুদ্ধার করা হয়।
স্মরণ করুন যে ATP এর ভাঙ্গন একটি এনজাইমেটিক হাইড্রোলাইসিস প্রতিক্রিয়া, এবং এটি সমীকরণ দ্বারা প্রকাশ করা যেতে পারে:
ATP-ase + ATP + H2O ---> ADP + H3PO4
ATP-এর পুনঃসংশ্লেষণের জন্য শক্তি (এটি বিভক্ত হওয়ার সময় প্রকাশ করা হবে - প্রায় 40 kJ প্রতি 1 mol) শক্তির মুক্তির (ক্যাটাবলিক) সাথে এগিয়ে যাওয়ার প্রতিক্রিয়াগুলির মাধ্যমে প্রাপ্ত করা আবশ্যক। অতএব, সেলুলার স্তরে, এটিপি হাইড্রোলাইসিসের প্রতিক্রিয়া এমন প্রতিক্রিয়াগুলির সাথে যুক্ত যা এটিপি পুনঃসংশ্লেষণ প্রদান করে। এই জাতীয় প্রতিক্রিয়াগুলির সময়, মধ্যবর্তী উচ্চ-শক্তি যৌগগুলি গঠিত হয় যার গঠনে একটি ফসফেট গ্রুপ থাকে, যা মুক্ত শক্তির রিজার্ভের সাথে ADP-তে স্থানান্তরিত হয়। ফসফোট্রান্সফেরেজ এনজাইম দ্বারা অনুঘটক করা এই ধরনের স্থানান্তর প্রতিক্রিয়া ("রিলে ব্যাটন" স্থানান্তর) ট্রান্সফসফোরিলেশন বা রিফসফোরিলেশন বিক্রিয়া বলে। এটিপি পুনঃসংশ্লেষণের জন্য প্রয়োজনীয় ম্যাক্রোঅার্জিক যৌগগুলি হয় ক্রমাগত উপস্থিত থাকে, উদাহরণস্বরূপ, ক্রিয়েটাইন ফসফেট (সিমপ্লাস্টে জমা হয়), বা অক্সিডেটিভ প্রক্রিয়ায় (ক্যাটাবলিক) গঠিত হয় (ডিফসফোগ্লিসারিক অ্যাসিড, ফসফোপাইরুভিক অ্যাসিড)।
পেশী ক্রিয়াকলাপের সময় এটিপি সংশ্লেষণ দুটি উপায়ে করা যেতে পারে: অক্সিজেনের অংশগ্রহণ ছাড়া প্রতিক্রিয়ার কারণে - অ্যানেরোবিক (যখন পেশীগুলিতে অক্সিজেন সরবরাহের সময় থাকে না বা কঠিন হয়) এবং কোষে অক্সিডেটিভ প্রক্রিয়ার কারণে (অক্সিজেনগুলির অংশগ্রহণের সাথে) অক্সিজেন যা আমরা শ্বাস নিই, এবং যা ক্রীড়াবিদ লোডের অধীনে দ্রুত শ্বাস নেয় এবং বিশ্রামের প্রাথমিক পর্যায়ে)।
মানুষের কঙ্কালের পেশীগুলিতে, তিন ধরণের অ্যানেরোবিক প্রক্রিয়া চিহ্নিত করা হয়েছে, যার সময় এটিপি পুনর্সংশ্লেষণ করা হয়:
- ক্রিয়েটাইন ফসফোকিনেস প্রতিক্রিয়া (ফসফোজেনিক বা অ্যালাকটিক অ্যানারোবিক প্রক্রিয়া), যেখানে ক্রিয়েটাইন ফসফেট এবং এডিপি-র মধ্যে রিফসফোরিলেশনের কারণে এটিপি পুনঃসংশ্লেষণ ঘটে;
- গ্লাইকোলাইসিস (ল্যাকট্যাসিড অ্যানেরোবিক প্রক্রিয়া), যেখানে এটিপি পুনঃসংশ্লেষণ করা হয় কার্বোহাইড্রেটের এনজাইমেটিক অ্যানেরোবিক ভাঙ্গনের সময়, ল্যাকটিক অ্যাসিড গঠনের সাথে শেষ হয়।
- মায়োকিনেস প্রতিক্রিয়া, যেখানে ADP-এর একটি নির্দিষ্ট অংশের dephosphorylation এর কারণে ATP resynthesis করা হয়;
পেশী ক্রিয়াকলাপের সময় বিভিন্ন ধরণের শক্তি রূপান্তর প্রক্রিয়াগুলির তুলনা এবং পরিমাপ করতে, তিনটি প্রধান মানদণ্ড ব্যবহার করা হয়:
- ক্ষমতার মানদণ্ড -একটি প্রদত্ত প্রক্রিয়ায় শক্তি রূপান্তরের হার নির্দেশ করে (ব্যায়াম);
- ক্ষমতার মানদণ্ড -শক্তি পদার্থের মোট মজুদ প্রতিফলিত করে (নিঃসৃত শক্তি এবং কাজ সম্পাদিত পরিমাণ দ্বারা পরিমাপ করা হয়);
- দক্ষতার মানদণ্ড - ATP পুনঃসংশ্লেষণে ব্যয় করা শক্তি এবং এই প্রক্রিয়া (ব্যায়াম) চলাকালীন মোট শক্তির পরিমাণের মধ্যে অনুপাত চিহ্নিত করে।
শক্তি রূপান্তর প্রক্রিয়া, অ্যানেরোবিক এবং বায়বীয়, শক্তি, ক্ষমতা এবং দক্ষতার মধ্যে পার্থক্য। উচ্চ তীব্রতার স্বল্পমেয়াদী অনুশীলনের সময় অ্যানেরোবিক প্রক্রিয়াগুলি প্রাধান্য পায়, বায়বীয় প্রক্রিয়াগুলি - মাঝারি তীব্রতার দীর্ঘমেয়াদী কাজের সময়।
বক্তৃতার সারাংশ| বক্তৃতার সারাংশ | ইন্টারেক্টিভ পরীক্ষা | বিমূর্ত ডাউনলোড করুন
» কঙ্কালের পেশীর কাঠামোগত সংগঠন
» কঙ্কালের পেশী সংকোচনের আণবিক প্রক্রিয়া
» কঙ্কালের পেশীতে উত্তেজনা এবং সংকোচনের সংযোগ
»কঙ্কালের পেশী শিথিলকরণ
»
» কঙ্কালের পেশীর কাজ
» মসৃণ পেশীগুলির কাঠামোগত সংগঠন এবং সংকোচন
»পেশীর শারীরবৃত্তীয় বৈশিষ্ট্য
পেশী সংকোচন শরীরের একটি গুরুত্বপূর্ণ কাজ যা প্রতিরক্ষামূলক, শ্বাসযন্ত্র, পুষ্টিকর, যৌন, রেচন এবং অন্যান্য শারীরবৃত্তীয় প্রক্রিয়াগুলির সাথে যুক্ত। সমস্ত ধরণের স্বেচ্ছাসেবী নড়াচড়া - হাঁটা, মুখের অভিব্যক্তি, চোখের গোলাগুলির নড়াচড়া, গিলে ফেলা, শ্বাস প্রশ্বাস ইত্যাদি কঙ্কালের পেশী দ্বারা সঞ্চালিত হয়। অনৈচ্ছিক নড়াচড়া (হার্টের সংকোচন ব্যতীত) - পাকস্থলী এবং অন্ত্রের পেরিস্টালসিস, রক্তনালীগুলির স্বরে পরিবর্তন, মূত্রাশয়ের স্বর বজায় রাখা - মসৃণ পেশীগুলির সংকোচনের কারণে ঘটে। হার্টের কাজ কার্ডিয়াক পেশীগুলির সংকোচনের দ্বারা সরবরাহ করা হয়।
কঙ্কালের পেশীগুলির কাঠামোগত সংগঠন
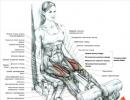
পেশী ফাইবার এবং মায়োফাইব্রিল (চিত্র 1)।কঙ্কালের পেশী অনেকগুলি পেশী তন্তু নিয়ে গঠিত যেগুলির হাড়ের সাথে সংযুক্তির বিন্দু রয়েছে এবং একে অপরের সমান্তরাল। প্রতিটি পেশী ফাইবার (মায়োসাইট) অনেকগুলি সাবুনিট অন্তর্ভুক্ত করে - মায়োফাইব্রিলস, যা অনুদৈর্ঘ্যভাবে পুনরাবৃত্তি করা ব্লক (সারকোমেরেস) থেকে তৈরি করা হয়। সারকোমের হল কঙ্কালের পেশীর সংকোচনযন্ত্রের কার্যকরী একক। পেশী ফাইবারে মায়োফাইব্রিলগুলি এমনভাবে থাকে যে তাদের মধ্যে সারকোমেরের অবস্থান মিলে যায়। এটি ট্রান্সভার্স স্ট্রিয়েশনের একটি প্যাটার্ন তৈরি করে।
সারকোমের এবং ফিলামেন্ট।মায়োফাইব্রিলের সারকোমেরেস জেড-প্লেট দ্বারা একে অপরের থেকে পৃথক করা হয়, যাতে প্রোটিন বিটা-অ্যাক্টিনিন থাকে। উভয় দিকে, পাতলা অ্যাক্টিন ফিলামেন্টগুলি জেড-প্লেট থেকে প্রসারিত হয়। তাদের মধ্যে ঘন মায়োসিন ফিলামেন্ট রয়েছে।
অ্যাক্টিন ফিলামেন্ট দেখতে দুটি স্ট্র্যান্ডের পুঁতির মতো একটি ডাবল হেলিক্সে পেঁচানো, যেখানে প্রতিটি পুঁতি একটি অ্যাক্টিন প্রোটিন অণু। অ্যাক্টিন হেলিসের অবকাশগুলিতে, একে অপরের থেকে সমান দূরত্বে, ফিলামেন্টাস ট্রপোমায়োসিন প্রোটিন অণুর সাথে সংযুক্ত ট্রপোনিন প্রোটিন অণু রয়েছে।
মায়োসিন ফিলামেন্টগুলি পুনরাবৃত্তি করা মায়োসিন প্রোটিন অণু দ্বারা গঠিত। প্রতিটি মায়োসিন অণুর একটি মাথা এবং একটি লেজ থাকে। মায়োসিন হেড অ্যাক্টিন অণুর সাথে আবদ্ধ হতে পারে, একটি তথাকথিত ক্রস ব্রিজ তৈরি করে।
পেশী ফাইবারের কোষের ঝিল্লি ইনভেজিনেশন (ট্রান্সভার্স টিউবুলস) গঠন করে, যা সারকোপ্লাজমিক রেটিকুলামের ঝিল্লিতে উত্তেজনা সঞ্চালনের কাজ করে। সারকোপ্লাজমিক রেটিকুলাম (লংগিটিউডিনাল টিউবুলস) হল বন্ধ টিউবুলের একটি আন্তঃকোষীয় নেটওয়ার্ক এবং এটি Ca ++ আয়ন জমা করার কাজ করে।
মোটর ইউনিট।কঙ্কাল পেশীর কার্যকরী একক হল মোটর ইউনিট (MU)। DE - পেশী তন্তুগুলির একটি সেট যা একটি মোটর নিউরনের প্রক্রিয়া দ্বারা উদ্ভূত হয়। এক এমইউ তৈরি করা ফাইবারগুলির উত্তেজনা এবং সংকোচন একই সাথে ঘটে (যখন সংশ্লিষ্ট মোটর নিউরন উত্তেজিত হয়)। স্বতন্ত্র MUs একে অপরের থেকে স্বাধীনভাবে ফায়ার এবং সংকোচন করতে পারে।
সংকোচনের আণবিক প্রক্রিয়াকঙ্কাল পেশী
ফিলামেন্ট স্লাইডিং তত্ত্ব অনুসারে, একে অপরের সাপেক্ষে অ্যাক্টিন এবং মায়োসিন ফিলামেন্টের স্লাইডিং আন্দোলনের কারণে পেশী সংকোচন ঘটে। থ্রেড স্লাইডিং মেকানিজম বেশ কয়েকটি ধারাবাহিক ইভেন্ট অন্তর্ভুক্ত করে।
মায়োসিন হেড অ্যাক্টিন ফিলামেন্ট বাইন্ডিং সাইটগুলির সাথে সংযুক্ত করে (চিত্র 2, এ)।
অ্যাক্টিনের সাথে মায়োসিনের মিথস্ক্রিয়া মায়োসিন অণুর গঠনমূলক পুনর্বিন্যাসের দিকে পরিচালিত করে। মাথা ATPase কার্যকলাপ অর্জন করে এবং 120° ঘোরায়। মাথার ঘূর্ণনের কারণে, অ্যাক্টিন এবং মায়োসিন ফিলামেন্ট একে অপরের সাপেক্ষে "এক ধাপ" সরে যায় (চিত্র 2বি)।
অ্যাক্টিন এবং মায়োসিনের বিচ্ছেদ এবং মাথার গঠন পুনরুদ্ধার ঘটে মায়োসিন মাথার সাথে একটি ATP অণুর সংযুক্তির ফলে এবং Ca++ (চিত্র 2, C) এর উপস্থিতিতে এর হাইড্রোলাইসিস।
চক্রটি "আবদ্ধকরণ - গঠনে পরিবর্তন - সংযোগ বিচ্ছিন্নতা - গঠনের পুনরুদ্ধার" অনেকবার ঘটে, যার ফলস্বরূপ অ্যাক্টিন এবং মায়োসিন ফিলামেন্টগুলি একে অপরের সাপেক্ষে স্থানচ্যুত হয়, সারকোমেরেসের জেড-ডিস্কগুলি একে অপরের কাছে আসে এবং মায়োফাইব্রিল ছোট হয় (চিত্র। 2, ডি)।
উত্তেজনা এবং সংকোচনের সংযোগকঙ্কালের পেশীতে
বিশ্রামে, মায়োফাইব্রিলে ফিলামেন্ট স্লাইডিং ঘটে না, যেহেতু অ্যাক্টিন পৃষ্ঠের বাঁধাই কেন্দ্রগুলি ট্রপোমায়োসিন প্রোটিন অণু দ্বারা বন্ধ থাকে (চিত্র 3, এ, বি)। মায়োফাইব্রিলগুলির উত্তেজনা (বিধ্বংসীকরণ) এবং সঠিক পেশী সংকোচন ইলেক্ট্রোমেকানিকাল কাপলিং প্রক্রিয়ার সাথে যুক্ত, যার মধ্যে বেশ কয়েকটি ধারাবাহিক ঘটনা রয়েছে।
পোস্টসিন্যাপটিক ঝিল্লিতে নিউরোমাসকুলার সিন্যাপস ফায়ারিংয়ের ফলে, একটি EPSP ঘটে, যা পোস্টসিন্যাপটিক ঝিল্লির আশেপাশের এলাকায় একটি অ্যাকশন পটেনশিয়াল তৈরি করে।
উত্তেজনা (অ্যাকশন পটেনশিয়াল) মায়োফাইব্রিল ঝিল্লি বরাবর ছড়িয়ে পড়ে এবং ট্রান্সভার্স টিউবুলের সিস্টেমের কারণে সারকোপ্লাজমিক রেটিকুলামে পৌঁছে। সারকোপ্লাজমিক রেটিকুলাম ঝিল্লির ডিপোলারাইজেশন এটিতে Ca++ চ্যানেল খোলার দিকে নিয়ে যায়, যার মাধ্যমে Ca++ আয়ন সারকোপ্লাজমে প্রবেশ করে (চিত্র 3, সি)।
Ca++ আয়ন ট্রপোনিন প্রোটিনের সাথে আবদ্ধ হয়। ট্রোপোনিন তার গঠন পরিবর্তন করে এবং ট্রপোমায়োসিন প্রোটিন অণুগুলিকে স্থানচ্যুত করে যা অ্যাক্টিন বাঁধাই কেন্দ্রগুলিকে বন্ধ করে দেয় (চিত্র 3d)।
মায়োসিন মাথা খোলা বাঁধাই কেন্দ্রে যোগদান করে, এবং সংকোচনের প্রক্রিয়া শুরু হয় (চিত্র 3, ই)।
এই প্রক্রিয়াগুলির বিকাশের জন্য, একটি নির্দিষ্ট সময়কাল (10-20 ms) প্রয়োজন। পেশী ফাইবার (পেশী) উত্তেজনার মুহূর্ত থেকে তার সংকোচনের শুরু পর্যন্ত সময়কে সংকোচনের সুপ্ত সময় বলা হয়।
কঙ্কালের পেশী শিথিলকরণ
সারকোপ্লাজমিক রেটিকুলামের চ্যানেলে ক্যালসিয়াম পাম্পের মাধ্যমে Ca++ আয়ন বিপরীত স্থানান্তরের কারণে পেশী শিথিল হয়। যেহেতু Ca++ সাইটোপ্লাজম থেকে সরানো হয়, সেখানে কম এবং কম খোলা বাঁধাই সাইট রয়েছে এবং অবশেষে অ্যাক্টিন এবং মায়োসিন ফিলামেন্টগুলি সম্পূর্ণরূপে মিলিত হয়; পেশী শিথিলতা ঘটে।
একটি সংকোচন হল একটি পেশীর ক্রমাগত দীর্ঘায়িত সংকোচন যা উদ্দীপনা বন্ধ হওয়ার পরেও অব্যাহত থাকে। সারকোপ্লাজমে প্রচুর পরিমাণে Ca ++ জমা হওয়ার ফলে টিটানিক সংকোচনের পরে স্বল্পমেয়াদী সংকোচনের বিকাশ হতে পারে; দীর্ঘমেয়াদী (কখনও কখনও অপরিবর্তনীয়) সংকোচন বিষক্রিয়া, বিপাকীয় ব্যাধির ফলে ঘটতে পারে।
কঙ্কালের পেশী সংকোচনের পর্যায় এবং মোড
পেশী সংকোচনের পর্যায়গুলি
যখন একটি কঙ্কালের পেশী সুপারথ্রেশহোল্ড শক্তির বৈদ্যুতিক প্রবাহের একক আবেগ দ্বারা উদ্দীপিত হয়, তখন একটি একক পেশী সংকোচন ঘটে, যার মধ্যে 3টি পর্যায় আলাদা করা হয় (চিত্র 4, এ):
সংকোচনের সুপ্ত (লুকানো) সময়কাল (প্রায় 10 এমএস), যে সময়ে অ্যাকশন পটেনশিয়াল বিকশিত হয় এবং ইলেক্ট্রোমেকানিকাল কাপলিং প্রক্রিয়াগুলি ঘটে; একক সংকোচনের সময় পেশীর উত্তেজনা কর্ম সম্ভাবনার পর্যায় অনুসারে পরিবর্তিত হয়;
সংক্ষিপ্তকরণ পর্ব (প্রায় 50 এমএস);
শিথিলকরণ পর্ব (প্রায় 50 এমএস)।
পেশী সংকোচনের মোড
প্রাকৃতিক অবস্থার অধীনে, শরীরের একটি একক পেশী সংকোচন পরিলক্ষিত হয় না, যেহেতু ক্রিয়া ক্ষমতার একটি সিরিজ মোটর স্নায়ুর সাথে যায় যা পেশীকে উদ্বুদ্ধ করে। পেশীতে স্নায়ু প্রবণতার কম্পাঙ্কের উপর নির্ভর করে, পেশী তিনটি মোডের একটিতে সংকোচন করতে পারে (চিত্র 4বি)।
একক পেশী সংকোচন বৈদ্যুতিক আবেগের কম ফ্রিকোয়েন্সিতে ঘটে। শিথিলকরণ পর্বের সমাপ্তির পর যদি পরবর্তী আবেগ পেশীতে আসে, তাহলে ধারাবাহিক একক সংকোচনের একটি সিরিজ ঘটে।
আবেগের উচ্চ কম্পাঙ্কে, পরবর্তী আবেগ পূর্ববর্তী সংকোচন চক্রের শিথিলকরণ পর্যায়ের সাথে মিলে যেতে পারে। সংকোচনের প্রশস্ততা সংক্ষিপ্ত করা হবে, একটি জ্যাগড টিটেনাস থাকবে - একটি দীর্ঘ সংকোচন, পেশীর অসম্পূর্ণ শিথিলতার সময়সীমার দ্বারা বাধাপ্রাপ্ত।
আবেগের ফ্রিকোয়েন্সি আরও বৃদ্ধির সাথে, প্রতিটি পরবর্তী আবেগ সংক্ষিপ্ত হওয়ার পর্যায়ে পেশীতে কাজ করবে, যার ফলে একটি মসৃণ টিটেনাস হবে - একটি দীর্ঘ সংকোচন যা শিথিলকরণের সময় বাধাগ্রস্ত হয় না।
ফ্রিকোয়েন্সি অপ্টিমাম এবং পেসিমাম
টিটানিক সংকোচনের প্রশস্ততা পেশীকে জ্বালাতনকারী আবেগের ফ্রিকোয়েন্সির উপর নির্ভর করে। ফ্রিকোয়েন্সি সর্বোত্তম হল বিরক্তিকর আবেগের এমন একটি ফ্রিকোয়েন্সি, যেখানে প্রতিটি পরবর্তী আবেগ বর্ধিত উত্তেজনার পর্যায়ের সাথে মিলে যায় (চিত্র 4, এ) এবং সেই অনুযায়ী, সর্বোচ্চ প্রশস্ততার টিটেনাস ঘটায়। একটি ফ্রিকোয়েন্সি পেসিমাম হল একটি উচ্চতর উদ্দীপনা ফ্রিকোয়েন্সি, যেখানে প্রতিটি পরবর্তী কারেন্ট পালস অবাধ্যতা পর্যায়ে পড়ে (চিত্র 4, এ), যার ফলস্বরূপ টিটেনাস প্রশস্ততা উল্লেখযোগ্যভাবে হ্রাস পায়।
কঙ্কাল পেশী কাজ
কঙ্কালের পেশী সংকোচনের শক্তি 2 টি কারণ দ্বারা নির্ধারিত হয়:
হ্রাসে অংশগ্রহণকারী এমইউ-এর সংখ্যা;
পেশী তন্তুগুলির সংকোচনের ফ্রিকোয়েন্সি।
কঙ্কালের পেশীর কাজটি সংকোচনের সময় পেশীর টোন (টেনশন) এবং দৈর্ঘ্যের একটি সমন্বিত পরিবর্তন দ্বারা সম্পন্ন হয়।
কঙ্কালের পেশীর কাজের ধরন:
গতিশীল ওভারকামিং কাজ সঞ্চালিত হয় যখন পেশী, সংকোচন, শরীর বা এর অংশগুলিকে স্থানান্তরিত করে;
স্থির (ধারণ) কাজ করা হয় যদি, পেশী সংকোচনের কারণে, শরীরের অংশগুলি একটি নির্দিষ্ট অবস্থানে রাখা হয়;
গতিশীল নিকৃষ্ট কাজ সঞ্চালিত হয় যদি পেশী কাজ করে, তবে একই সময়ে প্রসারিত হয়, যেহেতু এটি যে শক্তি তৈরি করে তা শরীরের অংশগুলি সরাতে বা ধরে রাখার জন্য যথেষ্ট নয়।
কাজের পারফরম্যান্সের সময়, পেশী সংকোচন করতে পারে:
আইসোটোনিক - ধ্রুবক টান (বাহ্যিক লোড) সহ পেশী ছোট হয়; আইসোটোনিক সংকোচন শুধুমাত্র পরীক্ষায় পুনরুত্পাদিত হয়;
আইসোমেট্রিক - পেশী টান বৃদ্ধি পায়, তবে এর দৈর্ঘ্য পরিবর্তন হয় না; স্থির কাজ সম্পাদন করার সময় পেশীগুলি আইসোমেট্রিকভাবে সংকুচিত হয়;
অক্সোটোনিক - পেশী টান ছোট হওয়ার সাথে সাথে পরিবর্তিত হয়; অক্সোটোনিক সংকোচন গতিশীল ওভারকামিং কাজের সময় সঞ্চালিত হয়।
গড় লোড নিয়ম- পেশী মাঝারি লোডের সাথে সর্বাধিক কাজ করতে পারে।
ক্লান্তি হল একটি পেশীর একটি শারীরবৃত্তীয় অবস্থা যা দীর্ঘ পরিশ্রমের পরে বিকাশ লাভ করে এবং সংকোচনের প্রশস্ততা হ্রাস, সংকোচনের সুপ্ত সময়ের বর্ধিতকরণ এবং শিথিলকরণ পর্যায়ের দ্বারা প্রকাশিত হয়। ক্লান্তির কারণগুলি হল: এটিপি হ্রাস, পেশীতে বিপাকীয় পণ্য জমে। ছন্দবদ্ধ কাজের সময় পেশীর ক্লান্তি সিন্যাপস ক্লান্তির চেয়ে কম। অতএব, যখন শরীর পেশীবহুল কাজ করে, তখন ক্লান্তি প্রাথমিকভাবে সিএনএস সিন্যাপসেস এবং নিউরোমাসকুলার সিন্যাপসের স্তরে বিকাশ লাভ করে।
কাঠামোগত সংগঠন এবং হ্রাসমসৃণ পেশী
কাঠামোগত সংগঠন। মসৃণ পেশী একক টাকু-আকৃতির কোষ (মায়োসাইট) নিয়ে গঠিত যা পেশীতে কমবেশি এলোমেলোভাবে অবস্থিত। সংকোচনশীল ফিলামেন্টগুলি অনিয়মিতভাবে সাজানো হয়, যার ফলস্বরূপ পেশীর কোন ট্রান্সভার্স স্ট্রিয়েশন নেই।
সংকোচনের প্রক্রিয়াটি কঙ্কালের পেশীর মতোই, তবে ফিলামেন্ট স্লাইডিংয়ের হার এবং এটিপি হাইড্রোলাইসিসের হার কঙ্কালের পেশীর তুলনায় 100-1000 গুণ কম।
উত্তেজনা এবং সংকোচনের সংযোগের প্রক্রিয়া। যখন একটি কোষ উত্তেজিত হয়, তখন Ca++ শুধুমাত্র সারকোপ্লাজমিক রেটিকুলাম থেকে নয়, আন্তঃকোষীয় স্থান থেকেও মায়োসাইটের সাইটোপ্লাজমে প্রবেশ করে। Ca++ আয়ন, ক্যালমোডুলিন প্রোটিনের অংশগ্রহণে, একটি এনজাইম (মায়োসিন কিনেস) সক্রিয় করে, যা ফসফেট গ্রুপকে এটিপি থেকে মায়োসিনে স্থানান্তর করে। ফসফরিলেটেড মায়োসিন হেড অ্যাক্টিন ফিলামেন্টের সাথে সংযুক্ত করার ক্ষমতা অর্জন করে।
মসৃণ পেশীগুলির সংকোচন এবং শিথিলকরণ। সারকোপ্লাজম থেকে Ca ++ আয়ন অপসারণের হার কঙ্কালের পেশীর তুলনায় অনেক কম, যার ফলে শিথিলতা খুব ধীরে ধীরে ঘটে। মসৃণ পেশী দীর্ঘ টনিক সংকোচন এবং ধীর ছন্দময় আন্দোলন করে। এটিপি হাইড্রোলাইসিসের কম তীব্রতার কারণে, মসৃণ পেশীগুলি দীর্ঘমেয়াদী সংকোচনের জন্য সর্বোত্তমভাবে অভিযোজিত হয়, যা ক্লান্তি এবং উচ্চ শক্তি খরচের দিকে পরিচালিত করে না।
পেশীগুলির শারীরবৃত্তীয় বৈশিষ্ট্য
কঙ্কাল এবং মসৃণ পেশীগুলির সাধারণ শারীরবৃত্তীয় বৈশিষ্ট্যগুলি হল উত্তেজনা এবং সংকোচনশীলতা। কঙ্কাল এবং মসৃণ পেশীগুলির তুলনামূলক বৈশিষ্ট্যগুলি টেবিলে দেওয়া হয়েছে। 6.1। কার্ডিয়াক পেশীগুলির শারীরবৃত্তীয় বৈশিষ্ট্য এবং বৈশিষ্ট্যগুলি "হোমিওস্ট্যাসিসের শারীরবৃত্তীয় প্রক্রিয়া" বিভাগে আলোচনা করা হয়েছে।
সারণি 7.1. কঙ্কাল এবং মসৃণ পেশীগুলির তুলনামূলক বৈশিষ্ট্য
|
সম্পত্তি |
কঙ্কাল পেশী |
মসৃণ পেশী |
|
ডিপোলারাইজেশন হার |
ধীর |
|
|
অবাধ্য সময়ের |
সংক্ষিপ্ত |
দীর্ঘ |
|
হ্রাসের প্রকৃতি |
দ্রুত ফাসিক |
ধীর টনিক |
|
শক্তি খরচ |
||
|
প্লাস্টিক |
||
|
অটোমেশন |
||
|
পরিবাহিতা |
||
|
উদ্ভাবন |
সোমাটিক এনএসের মোটোনুরন |
স্বায়ত্তশাসিত NS এর পোস্টগ্যাংলিওনিক নিউরন |
|
আন্দোলন চালানো হয় |
ইচ্ছামত |
অনিচ্ছাকৃত |
|
রাসায়নিকের প্রতি সংবেদনশীলতা |
||
|
বিভাজন এবং পার্থক্য করার ক্ষমতা |
মসৃণ পেশীগুলির প্লাস্টিকতা এই সত্যে প্রকাশিত হয় যে তারা সংক্ষিপ্ত এবং প্রসারিত উভয় অবস্থায়ই একটি ধ্রুবক স্বন বজায় রাখতে পারে।
মসৃণ পেশী টিস্যুর পরিবাহিতা এই সত্যে প্রকাশিত হয় যে উত্তেজনা বিশেষ বৈদ্যুতিক পরিবাহী যোগাযোগের (নেক্সাস) মাধ্যমে এক মায়োসাইট থেকে অন্যটিতে ছড়িয়ে পড়ে।
মসৃণ পেশীগুলির স্বয়ংক্রিয়তার বৈশিষ্ট্যটি এই সত্যে প্রকাশিত হয় যে এটি স্নায়ুতন্ত্রের অংশগ্রহণ ছাড়াই সংকোচন করতে পারে, কারণ কিছু মায়োসাইট স্বতঃস্ফূর্তভাবে ছন্দময়ভাবে পুনরাবৃত্তিমূলক কর্ম সম্ভাবনা তৈরি করতে সক্ষম হয়।
শরীরের সমস্ত পেশী মসৃণ এবং স্ট্রাইটেড বিভক্ত।
কঙ্কালের পেশী সংকোচনের প্রক্রিয়া
স্ট্রিয়েটেড পেশী দুটি প্রকারে বিভক্ত: কঙ্কালের পেশী এবং মায়োকার্ডিয়াম।
পেশী ফাইবারের গঠন
পেশী কোষের ঝিল্লি, যাকে সারকোলেমা বলা হয়, বৈদ্যুতিকভাবে উত্তেজনাপূর্ণ এবং একটি কর্ম সম্ভাবনা পরিচালনা করতে সক্ষম। পেশী কোষে এই প্রক্রিয়াগুলি স্নায়ু কোষের মতো একই নীতি অনুসারে ঘটে। পেশী ফাইবারের বিশ্রামের সম্ভাবনা প্রায় -90 mV, অর্থাৎ নার্ভ ফাইবারের (-70 mV) থেকে কম; ক্রিটিক্যাল ডিপোলারাইজেশন, যেখানে পৌঁছানোর পর একটি অ্যাকশন পটেনশিয়াল উদ্ভূত হয়, তা স্নায়ু তন্তুর মতোই। তাই: পেশী ফাইবারের উত্তেজনা স্নায়ু ফাইবারের উত্তেজনার চেয়ে কিছুটা কম, যেহেতু পেশী কোষকে প্রচুর পরিমাণে বিধ্বংসী হতে হবে।
উদ্দীপনা একটি পেশী ফাইবার প্রতিক্রিয়া হয় হ্রাস, যা কোষের সংকোচন যন্ত্র তৈরি করে - মায়োফাইব্রিলস. তারা দুই ধরনের থ্রেড গঠিত strands: পুরু - মায়োসিন, এবং পাতলা actin. পুরু ফিলামেন্টে (15 nm ব্যাস এবং 1.5 µm লম্বা) শুধুমাত্র একটি প্রোটিন, মায়োসিন থাকে। পাতলা ফিলামেন্টে (7 nm ব্যাস এবং 1 µm লম্বা) তিন ধরনের প্রোটিন থাকে: অ্যাক্টিন, ট্রপোমায়োসিন এবং ট্রপোনিন।
actinএটি একটি দীর্ঘ প্রোটিন ফিলামেন্ট, যা পৃথক গ্লোবুলার প্রোটিন নিয়ে এমনভাবে সংযুক্ত থাকে যে পুরো কাঠামোটি একটি প্রসারিত চেইন। গ্লোবুলার অ্যাক্টিন (জি-অ্যাক্টিন) এর অণুগুলির অন্যান্য অনুরূপ অণুর সাথে পার্শ্বীয় এবং টার্মিনাল বাইন্ডিং সাইট রয়েছে। ফলস্বরূপ, তারা এমনভাবে একত্রিত হয় যে তারা একটি কাঠামো তৈরি করে যা প্রায়শই একসাথে সংযুক্ত দুটি পুঁতির সাথে তুলনা করা হয়। জি-অ্যাক্টিন অণু থেকে গঠিত পটি একটি সর্পিল মধ্যে পেঁচানো হয়। এই গঠনকে বলা হয় ফাইব্রিলার অ্যাক্টিন (এফ-অ্যাক্টিন)। হেলিক্স পিচ (কুণ্ডলীর দৈর্ঘ্য) 38 এনএম; প্রতিটি হেলিক্স কয়েলে 7 জোড়া জি-অ্যাক্টিন রয়েছে। জি-অ্যাক্টিনের পলিমারাইজেশন, অর্থাৎ, এফ-অ্যাক্টিনের গঠন, এটিপি-এর শক্তির কারণে ঘটে এবং বিপরীতভাবে, যখন এফ-অ্যাক্টিন ধ্বংস হয়ে যায়, তখন শক্তি নির্গত হয়।

আকার 1. এফ-অ্যাকটিনে পৃথক জি-অ্যাক্টিন গ্লোবুলসের ফিউশন
অ্যাক্টিন ফিলামেন্টের সর্পিল খাঁজ বরাবর প্রোটিন ট্রপোমায়োসিন রয়েছে। ট্রপোমায়োসিনের প্রতিটি স্ট্র্যান্ড, যা 41 এনএম দীর্ঘ, দুটি অভিন্ন α-শৃঙ্খল নিয়ে গঠিত, 7 এনএম দৈর্ঘ্যের বাঁক নিয়ে একত্রে পেঁচানো হয়। ট্রপোমায়োসিনের দুটি অণু এফ-অ্যাক্টিনের এক বাঁক বরাবর অবস্থিত। প্রতিটি ট্রপোমায়োসিন অণু পরেরটির সাথে সামান্য ওভারল্যাপ করে, ফলে একটি ট্রপোমায়োসিন ফিলামেন্ট তৈরি হয় যা অ্যাক্টিন বরাবর ক্রমাগত প্রসারিত হয়।

চিত্র 2। মায়োফাইব্রিলের একটি পাতলা ফিলামেন্টের গঠন
স্ট্রাইটেড পেশী কোষগুলিতে, অ্যাক্টিন এবং ট্রপোমায়োসিন ছাড়াও, পাতলা ফিলামেন্টের সংমিশ্রণে প্রোটিন ট্রপোনিনও অন্তর্ভুক্ত থাকে। এই গ্লোবুলার প্রোটিনের একটি জটিল গঠন রয়েছে। এটি তিনটি সাবইউনিট নিয়ে গঠিত, যার প্রতিটি সংকোচন প্রক্রিয়ায় তার কার্য সম্পাদন করে।
পুরু থ্রেডঅনেক অণু দ্বারা গঠিত মায়োসিনএকটি বান্ডিল মধ্যে সংগৃহীত। 155 এনএম লম্বা এবং 2 এনএম ব্যাসের প্রতিটি মায়োসিন অণু ছয়টি পলিপেপটাইড ফিলামেন্ট নিয়ে গঠিত: দুটি দীর্ঘ এবং চারটি ছোট। লম্বা চেইনগুলি 7.5 এনএম পিচ হেলিক্সে একত্রে কুণ্ডলী করা হয় এবং মায়োসিন অণুর ফাইব্রিলার অংশ গঠন করে। অণুর এক প্রান্তে, এই চেইনগুলি খুলে যায় এবং একটি কাঁটাযুক্ত প্রান্ত তৈরি করে। এই প্রান্তগুলির প্রতিটি দুটি ছোট চেইন সহ একটি জটিল গঠন করে, অর্থাৎ প্রতিটি অণুর উপরে দুটি মাথা রয়েছে। এটি মায়োসিন অণুর গ্লোবুলার অংশ।

চিত্র 3. মায়োসিন অণুর গঠন।
মায়োসিনে দুটি খণ্ডকে আলাদা করা হয়েছে: হালকা মেরোমায়োসিন (এলএমএম) এবং ভারী মেরোমায়োসিন (এইচএমএম), তাদের মধ্যে একটি কবজা রয়েছে। TMM দুটি সাবফ্র্যাগমেন্ট নিয়ে গঠিত: S1 এবং S2। LMM এবং সাবফ্র্যাগমেন্ট S2 থ্রেডের একটি বান্ডিলে নেস্ট করা হয় এবং সাবফ্র্যাগমেন্ট S1 পৃষ্ঠের উপরে প্রসারিত হয়। এই প্রসারিত প্রান্ত (মায়োসিন হেড) অ্যাক্টিন ফিলামেন্টের সক্রিয় সাইটে আবদ্ধ হতে এবং মায়োসিন ফিলামেন্টের বান্ডিলের দিকে ঝোঁকের কোণ পরিবর্তন করতে সক্ষম। এলএমএমগুলির মধ্যে ইলেক্ট্রোস্ট্যাটিক মিথস্ক্রিয়াগুলির কারণে একটি বান্ডিলে পৃথক মায়োসিন অণুর সংমিশ্রণ ঘটে। থ্রেডের কেন্দ্রীয় অংশে কোন মাথা নেই। মায়োসিন অণুর সমগ্র কমপ্লেক্স 1.5 µm এর বেশি বিস্তৃত। এটি প্রকৃতিতে পরিচিত সবচেয়ে বড় জৈবিক আণবিক গঠনগুলির মধ্যে একটি।
একটি পোলারাইজিং মাইক্রোস্কোপের মাধ্যমে একটি স্ট্রেটেড পেশীর অনুদৈর্ঘ্য বিভাগ দেখার সময়, হালকা এবং অন্ধকার এলাকাগুলি দৃশ্যমান হয়। অন্ধকার অঞ্চলগুলি (ডিস্কগুলি) অ্যানিসোট্রপিক: মেরুকৃত আলোতে, তারা অনুদৈর্ঘ্য দিকে স্বচ্ছ এবং অনুপ্রস্থ দিকে অস্বচ্ছ দেখায়, A অক্ষর দ্বারা চিহ্নিত করা হয়। হালকা অঞ্চলগুলি আইসোট্রপিক এবং I অক্ষর দ্বারা চিহ্নিত করা হয়। ডিস্ক I শুধুমাত্র পাতলা ফিলামেন্টগুলি অন্তর্ভুক্ত করে , এবং ডিস্ক A - এবং পুরু এবং পাতলা। ডিস্ক A এর মাঝখানে, একটি হালকা ফালা দৃশ্যমান, যাকে H-জোন বলা হয়। এটিতে পাতলা থ্রেড নেই। ডিস্ক I একটি পাতলা স্ট্রিপ Z দ্বারা বিভক্ত, এটি একটি ঝিল্লি যাতে কাঠামোগত উপাদান থাকে যা পাতলা থ্রেডের প্রান্তগুলিকে একসাথে বেঁধে রাখে। দুটি জেড লাইনের মধ্যবর্তী এলাকাকে বলা হয় sarcomere.

চিত্র 4. মায়োফাইব্রিল গঠন (ক্রস সেকশন)

চিত্র.5। স্ট্রাইটেড পেশীর গঠন (অনুদৈর্ঘ্য বিভাগ)
প্রতিটি পুরু থ্রেড ছয়টি পাতলা দিয়ে ঘেরা, এবং প্রতিটি পাতলা সুতো তিনটি পুরু দ্বারা বেষ্টিত। এইভাবে, একটি ক্রস বিভাগে, পেশী ফাইবার একটি নিয়মিত ষড়ভুজাকার গঠন আছে।
পেশী সংকোচন
পেশী সংকোচনের সময়, অ্যাক্টিন এবং মায়োসিন ফিলামেন্টের দৈর্ঘ্য পরিবর্তিত হয় না। একে অপরের তুলনায় কেবল তাদের স্থানচ্যুতি রয়েছে: পাতলা থ্রেডগুলি পুরুগুলির মধ্যে ফাঁকে চলে যায়। এই ক্ষেত্রে, ডিস্ক A এর দৈর্ঘ্য অপরিবর্তিত থাকে এবং ডিস্ক I ছোট করে, স্ট্রিপ H প্রায় অদৃশ্য হয়ে যায়। পুরু এবং পাতলা ফিলামেন্টের মধ্যে ট্রান্সভার্স ব্রিজ (মায়োসিন হেড) থাকার কারণে এই ধরনের স্লাইডিং সম্ভব। সংকোচনের সাথে, সারকোমেরের দৈর্ঘ্য প্রায় 2.5 থেকে 1.7 মাইক্রন পর্যন্ত পরিবর্তন করা সম্ভব।
মায়োসিন ফিলামেন্টের অনেকগুলি মাথা রয়েছে যা দিয়ে এটি অ্যাক্টিনের সাথে আবদ্ধ হতে পারে। অ্যাক্টিন ফিলামেন্টে এমন সাইট (সক্রিয় কেন্দ্র) রয়েছে যেখানে মায়োসিন মাথা সংযুক্ত করতে পারে। একটি বিশ্রাম পেশী কোষে, এই বাঁধাই স্থানগুলি ট্রপোমায়োসিন অণু দ্বারা আবৃত থাকে, যা পাতলা এবং পুরু ফিলামেন্টের মধ্যে একটি বন্ধন গঠনে বাধা দেয়।
অ্যাক্টিন এবং মায়োসিন মিথস্ক্রিয়া করার জন্য, ক্যালসিয়াম আয়ন অবশ্যই উপস্থিত থাকতে হবে। বিশ্রামে, তারা সারকোপ্লাজমিক রেটিকুলামে অবস্থিত। এই অর্গানেল হল একটি ঝিল্লির গহ্বর যাতে একটি ক্যালসিয়াম পাম্প থাকে, যা ATP-এর শক্তি ব্যবহার করে ক্যালসিয়াম আয়নগুলিকে সারকোপ্লাজমিক রেটিকুলামে পরিবহন করে। এর অভ্যন্তরীণ পৃষ্ঠে Ca2+ আবদ্ধ করতে সক্ষম প্রোটিন রয়েছে, যা সাইটোপ্লাজম এবং রেটিকুলাম গহ্বরের মধ্যে এই আয়নগুলির ঘনত্বের পার্থক্য কিছুটা কমিয়ে দেয়। কোষের ঝিল্লি বরাবর প্রচারিত অ্যাকশন পটেনশিয়াল কোষের পৃষ্ঠের কাছাকাছি অবস্থিত রেটিকুলাম ঝিল্লিকে সক্রিয় করে এবং সাইটোপ্লাজমে Ca2+ নিঃসরণ ঘটায়।
ট্রপোনিন অণুর ক্যালসিয়ামের জন্য উচ্চ সম্পর্ক রয়েছে।
এর প্রভাবে, এটি ট্রপোমায়োসিন ফিলামেন্টের অবস্থানকে অ্যাক্টিন ফিলামেন্টে এমনভাবে পরিবর্তন করে যে সক্রিয় কেন্দ্রটি আগে ট্রপোমায়োসিন দ্বারা আবৃত ছিল, খুলে যায়। একটি ট্রান্সভার্স ব্রিজ খোলা সক্রিয় কেন্দ্রে যোগ দেয়। এটি মায়োসিনের সাথে অ্যাক্টিনের মিথস্ক্রিয়া ঘটায়। বন্ড গঠনের পর, মায়োসিন হেড, পূর্বে ফিলামেন্টের সমকোণে অবস্থিত ছিল, মায়োসিন হেডের সাপেক্ষে অ্যাক্টিন ফিলামেন্টকে প্রায় 10 এনএম করে কাত করে এবং টানে। গঠিত অ্যাটিন-মায়োসিন কমপ্লেক্স একে অপরের সাপেক্ষে থ্রেডগুলির আরও স্লাইডিং প্রতিরোধ করে, তাই এর বিচ্ছেদ প্রয়োজনীয়। এটি শুধুমাত্র ATP এর শক্তির কারণেই সম্ভব। মায়োসিনের ATPase কার্যকলাপ রয়েছে, অর্থাৎ এটি ATP হাইড্রোলাইসিস ঘটাতে সক্ষম। এই প্রক্রিয়ায় নির্গত শক্তি অ্যাক্টিন এবং মায়োসিনের মধ্যে বন্ধন ভেঙে দেয় এবং মায়োসিন হেড অ্যাক্টিন অণুর নতুন অংশের সাথে যোগাযোগ করতে সক্ষম হয়। সেতুগুলির কাজ এমনভাবে সিঙ্ক্রোনাইজ করা হয় যে এক সুতার সমস্ত সেতুর বাঁধন, কাত এবং ভাঙ্গা একই সাথে ঘটে। পেশী শিথিল হলে, ক্যালসিয়াম পাম্পের কাজ সক্রিয় হয়, যা সাইটোপ্লাজমে Ca2+ ঘনত্ব কমিয়ে দেয়; অতএব, পাতলা এবং পুরু সুতার মধ্যে বন্ধন আর গঠিত হতে পারে না। এই অবস্থার অধীনে, যখন প্রসারিত হয়, থ্রেডের পেশীগুলি একে অপরের সাথে অবাধে স্লাইড করে। যাইহোক, এই ধরনের এক্সটেনসিবিলিটি শুধুমাত্র এটিপির উপস্থিতিতেই সম্ভব। কোষে ATP না থাকলে অ্যাক্টিন-মায়োসিন কমপ্লেক্স ভেঙ্গে যেতে পারে না। থ্রেডগুলি একে অপরের সাথে কঠোরভাবে সংযুক্ত থাকে। এই ঘটনাটি কঠোর মরটিসে পরিলক্ষিত হয়।
Fig.6. সারকোমেরের হ্রাস: 1 - মায়োসিন থ্রেড; 2 - সক্রিয় কেন্দ্র; 3 - অ্যাক্টিন ফিলামেন্ট; 4 - মায়োসিন মাথা; 5 - জেড লাইন।
ক)পাতলা এবং পুরু থ্রেডের মধ্যে কোন মিথস্ক্রিয়া নেই;
খ) Ca2+ এর উপস্থিতিতে, মায়োসিন হেড অ্যাক্টিন ফিলামেন্টের সক্রিয় সাইটের সাথে আবদ্ধ হয়;

ভিতরে)ট্রান্সভার্স ব্রিজগুলি তুলনামূলকভাবে পুরু দিয়ে একটি পাতলা থ্রেড বাঁক করে এবং টান দেয়, যার ফলস্বরূপ সারকোমেরের দৈর্ঘ্য হ্রাস পায়;
ছ) ATP-এর শক্তির কারণে থ্রেডগুলির মধ্যে বন্ধনগুলি ভেঙে গেছে, মায়োসিনের মাথাগুলি নতুন সক্রিয় কেন্দ্রগুলির সাথে যোগাযোগ করতে প্রস্তুত।
পেশী সংকোচনের দুটি পদ্ধতি রয়েছে: আইসোটোনিক(ফাইবারের দৈর্ঘ্য পরিবর্তিত হয়, কিন্তু ভোল্টেজ অপরিবর্তিত থাকে) এবং আইসোমেট্রিক(পেশীর প্রান্তগুলি স্থিরভাবে স্থির থাকে, যার ফলস্বরূপ এটি পরিবর্তিত দৈর্ঘ্য নয়, তবে টান)।
পেশী সংকোচনের শক্তি এবং গতি
পেশীর গুরুত্বপূর্ণ বৈশিষ্ট্য হল শক্তি এবং সংকোচনের গতি। এই বৈশিষ্ট্যগুলি প্রকাশকারী সমীকরণগুলি পরীক্ষামূলকভাবে এ. হিল দ্বারা প্রাপ্ত হয়েছিল এবং পরবর্তীকালে পেশী সংকোচনের গতি তত্ত্ব (দেশেরেভস্কির মডেল) দ্বারা নিশ্চিত করা হয়েছিল।
পার্বত্য সমীকরণ, যা পেশী সংকোচনের শক্তি এবং গতির সাথে সম্পর্কিত, এর নিম্নলিখিত ফর্ম রয়েছে: (P+a)(v+b) = (P0+a)b = a(vmax+b), যেখানে v হল পেশী ছোট করার গতি; পি - পেশী বল বা এটি প্রয়োগ করা লোড; vmax হল পেশী ছোট করার সর্বোচ্চ গতি; P0 হল সংকোচনের আইসোমেট্রিক মোডে পেশী দ্বারা বিকশিত বল; a,b ধ্রুবক। সাধারণ ক্ষমতাপেশী দ্বারা বিকশিত সূত্র দ্বারা নির্ধারিত হয়: মোট = (P+a)v = b(P0-P). দক্ষতাপেশী স্থির থাকে ( প্রায় 40%) বল মানের পরিসরে 0.2 P0 থেকে 0.8 P0 পর্যন্ত। পেশী সংকোচনের সময়, একটি নির্দিষ্ট পরিমাণ তাপ নির্গত হয়। এই মান বলা হয় তাপ উৎপাদন. তাপ উত্পাদন শুধুমাত্র পেশী দৈর্ঘ্যের পরিবর্তনের উপর নির্ভর করে এবং লোডের উপর নির্ভর করে না। ধ্রুবক কএবং খএকটি প্রদত্ত পেশী জন্য ধ্রুবক মান আছে. ধ্রুবক কবল মাত্রা আছে, এবং খ- দ্রুততা. ধ্রুবক খতাপমাত্রার উপর অনেকাংশে নির্ভরশীল। ধ্রুবক ক 0.25 P0 থেকে 0.4 P0 পর্যন্ত মানের মধ্যে রয়েছে। এই তথ্যের ভিত্তিতে, এটি অনুমান করা হয় সর্বাধিক সংকোচনের গতিএই পেশী জন্য: vmax = b (P0/a).
পেশী টিস্যুর বৈশিষ্ট্য।
কঙ্কালের পেশী সংকোচন এবং এর প্রক্রিয়া
পেশী টিস্যুর প্রকারভেদ। অ্যাক্টিন-মায়োসিন কমপ্লেক্স এবং এর কার্যকারিতার প্রক্রিয়া।
3 ধরনের প্রাণী টিস্যু আছে: 1) পেশীবহুল, 2) স্নায়বিক, 3) গোপনীয়। প্রথমটি সংকোচন এবং আন্দোলনের কাজ বাস্তবায়নের মাধ্যমে উত্তেজনার প্রতিক্রিয়া জানায়। দ্বিতীয়টি - আবেগগুলি পরিচালনা এবং বিশ্লেষণ করার ক্ষমতা, তৃতীয়টি - বিভিন্ন গোপনীয়তা হাইলাইট করার জন্য।
পেশী টিস্যু 3 প্রকার: 1. স্ট্রিটেড, 2. মসৃণ, 3. কার্ডিয়াক।
| বৈশিষ্ট্য | striated | মসৃণ | কার্ডিয়াক |
| বিশেষীকরণ | সুউচ্চ | অন্তত বিশেষীকরণ। | মাধ্যমিক বিশেষীকরণ। |
| গঠন | 10 সেমি পর্যন্ত লম্বা ফাইবার, সাবুনিটে বিভক্ত - সারকোমেরেস। তন্তুগুলি সংযোগকারী টিস্যু, রক্তনালী দ্বারা আন্তঃসংযুক্ত। স্নায়ু শেষগুলি ফাইবারগুলির সাথে সংযোগ করে নিউরোমাসকুলার জংশন তৈরি করে। | পৃথক টাকু মত গঠিত. বান্ডিল মধ্যে সংযুক্ত কোষ. | প্রান্তের কোষগুলি শাখা থেকে বেরিয়ে আসে, প্রক্রিয়াগুলির সাহায্যে একে অপরের সাথে সংযোগ স্থাপন করে। |
| নিউক্লিয়াস | পেরিফেরিতে একাধিক কোর | কেন্দ্রে 1 কোর | কেন্দ্রে বেশ কয়েকটি কোর |
| সাইটোপ্লাজম | মাইটোকন্ড্রিয়া, সারকোপ্লাজম রয়েছে। রেটিকুলাম, টি টিউব, গ্লাইকোজেন, চর্বি ফোঁটা | সোড মাইটোকন্ড্রিয়ন, সারকোপ্লাজম। জালিকা, টিটিউব, | সোড মাইটোকন্ড্রিয়ন, সারকোপ্লাজম। রেটিকুলাম, টি টিউব, |
| sarcolemma | এখানে | না | এখানে |
| প্রবিধান | নিউরোজেনিক | নিউরোজেনিক | নিউরোগ এবং হাস্যকর |
| তির্যক ফিতে | এখানে | না | এখানে |
| সংযোগ কার্যকলাপ. | শক্তিশালী, দ্রুত সংকোচন। অবাধ্যতার সময়কাল সংক্ষিপ্ত - বিশ্রামের সময় কম। দ্রুত ক্লান্তি। | ধীর বীট | দ্রুত ছন্দ, দীর্ঘ অবাধ্য সময় - কোন ক্লান্তি নেই। |
অ্যাক্টিন-মায়োসিন কমপ্লেক্স।সমস্ত পেশী কোষ প্রচুর পরিমাণে বিশেষ সংকোচনযোগ্য প্রোটিন থাকে - এগুলি পেশী প্রোটিনের মোট পরিমাণের 60-80%। প্রধান সংকোচনশীল
প্রোটিন হল ফাইব্রিলার প্রোটিন: - মায়োসিন- পুরু থ্রেড গঠন করে; - actin- পাতলা থ্রেড গঠন করে। সংকোচন নিয়ন্ত্রণ করতে, গ্লোবুলার প্রোটিন ব্যবহার করা হয়: ট্রপোনিন-ট্রপোমায়োসিন।
মায়োসিন হল একটি 2-স্ট্র্যান্ড গঠন 1=180 nm এবং 0=2.5 nm। অ্যাক্টিন একটি 2-হেলিক্স পেপটাইড চেইন।
হ্রাস প্রক্রিয়া:ফাইব্রিলের অ্যাক্টিন এবং মায়োসিন স্থানিকভাবে পৃথক করা হয়। স্নায়ু প্ররোচনা নিউরোমাসকুলার জংশনের সিনাপটিক ফাটলে অ্যাসিটাইলকোলিন নিঃসরণ ঘটায়। এটা
নিউরোট্রান্সমিটার বাইন্ডিং এর পরে পোস্টসিন্যাপটিক ঝিল্লির ডিপোলারাইজেশন ঘটায় এবং
কোষের ঝিল্লি জুড়ে এবং পেশীতে কর্ম সম্ভাবনার প্রচার
টি টিউবের মাধ্যমে ফাইবার। অ্যাক্টিন-মায়োসিন মিথস্ক্রিয়ার ফলে, ফাইব্রিল সংকোচন ঘটে। এটি একটি সেতু গঠনের ফলে মায়োসিন মাথার সাথে অ্যাক্টিন ফিলামেন্টকে ধাক্কা দিয়ে অর্জন করা হয়। আবেগ অদৃশ্য হয়ে গেলে, Ca2+ পুনরুদ্ধার করা হয়, অ্যাক্টিন এবং মায়োসিনের মধ্যে সেতুটি ধ্বংস হয়ে যায় এবং পেশীটি তার আসল অবস্থায় ফিরে আসে।
ট্রোপোনিন হল একটি গ্লোবুলার প্রোটিন যার 3টি কেন্দ্র রয়েছে:
- টি - ট্রপোমায়োসিনের সাথে আবদ্ধ হয়
- C - Ca2 + আবদ্ধ করে
- 1 - অ্যাক্টিন-মায়োসিন মিথস্ক্রিয়াকে বাধা দেয়।
সংকোচন পর্যায়গুলি:
1. সুপ্ত সময়কাল - 0.05 সেকেন্ড।
2. সংকোচন পর্যায় - 0.1 সেকেন্ড
3. বিশ্রামের সময়কাল - 0.2 সেকেন্ড।
পেশী কাজের বায়োকেমিস্ট্রি
1. ATP + মায়োসিন-অ্যাক্টিন কমপ্লেক্স ——-ADP + মায়োসিন + অ্যাক্টিন + F + শক্তি
2. ADP + ক্রিয়েটিনাইন ফসফেট——ATP + ক্রিয়েটাইন
3. গ্লাইকোজেন-গ্লুকোজ--গ্লুকোজ + O2--CO2 + H2O + 38 ATP (বায়ুবিক প্রক্রিয়া)
4. গ্লুকোজ - 2 ল্যাকটিক অ্যাসিড + 2 ATP
5. মিল্ক অ্যাসিড + O2-CO2 + H2O (বিশ্রাম) বা Mol.k-ta-গ্লুকোজ-গ্লাইকোজেন।
কঙ্কালের পেশী সংকোচনের প্রক্রিয়া
পেশী ছোট হওয়া একাধিক সারকোমেরের সংকোচনের ফলাফল।যখন অ্যাক্টিন ফিলামেন্টগুলি ছোট করা হয়, তখন তারা মায়োসিন ফিলামেন্টের তুলনায় স্লাইড করে, যার ফলস্বরূপ পেশী ফাইবারের প্রতিটি সারকোমেরের দৈর্ঘ্য হ্রাস পায়। এই ক্ষেত্রে, থ্রেডগুলির দৈর্ঘ্য অপরিবর্তিত থাকে। মায়োসিন ফিলামেন্টের ট্রান্সভার্স প্রোট্রুশন (ক্রস ব্রিজ) প্রায় 20 এনএম লম্বা। প্রতিটি প্রোট্রুশন একটি মাথা নিয়ে গঠিত, যা একটি "ঘাড়" (চিত্র 23) এর মাধ্যমে মায়োসিন ফিলামেন্টের সাথে সংযুক্ত থাকে।
একটি শিথিল অবস্থায়, ট্রান্সভার্স ব্রিজগুলির মাথার পেশীগুলি অ্যাক্টিন ফিলামেন্টগুলির সাথে যোগাযোগ করতে পারে না, যেহেতু তাদের সক্রিয় সাইটগুলি (মাথার সাথে পারস্পরিক যোগাযোগের জায়গাগুলি) ট্রপোমায়োসিন দ্বারা বিচ্ছিন্ন হয়। পেশী সংক্ষিপ্ত হওয়া ট্রান্সভার্স ব্রিজের গঠনগত পরিবর্তনের ফলাফল: এর মাথা "ঘাড়" বাঁকিয়ে কাত হয়ে যায়।

ভাত। 23. স্ট্রাইটেড পেশীতে সংকোচনযোগ্য এবং নিয়ন্ত্রক প্রোটিনের স্থানিক সংগঠন। মায়োসিন সেতুর অবস্থান (স্ট্রোক প্রভাব, ঘাড় বাঁকানো) পেশী ফাইবার (ফাইবার সংকোচন) মধ্যে সংকোচনশীল প্রোটিনের মিথস্ক্রিয়া প্রক্রিয়াতে দেখানো হয়
প্রক্রিয়া ক্রম , প্রদান পেশী ফাইবার সংকোচন(ইলেক্ট্রোমেকানিকাল ইন্টারফেস):
1. ঘটনার পর পিডিসিনাপসের কাছাকাছি পেশী ফাইবারে (পিসিপির বৈদ্যুতিক ক্ষেত্রের কারণে) উত্তেজনা মায়োসাইট মেমব্রেন জুড়ে ছড়িয়ে পড়ে, অনুপ্রস্থের ঝিল্লি সহ টি-টিউবুলস. একটি পেশী ফাইবার বরাবর AP পরিবাহনের প্রক্রিয়াটি একটি অমিলিনেটেড নার্ভ ফাইবার বরাবরের মতোই - ফলে সিন্যাপসের কাছাকাছি AP, তার বৈদ্যুতিক ক্ষেত্রের মাধ্যমে, ফাইবারের সংলগ্ন অংশে নতুন AP-এর উত্থান নিশ্চিত করে। (উত্তেজনার ক্রমাগত সঞ্চালন)
2. সম্ভাব্যকর্ম টি-টিউবুলসএর বৈদ্যুতিক ক্ষেত্রের কারণে ভোল্টেজ-গেটেড ক্যালসিয়াম চ্যানেলগুলি সক্রিয় করে মেমব্রেন এসপিআর, যার ফলে Ca2+ইলেক্ট্রোকেমিক্যাল গ্রেডিয়েন্ট অনুসারে এসপিআর ট্যাঙ্কগুলি ছেড়ে যায়।
3. ইন্টারফিব্রিলার স্পেসে Ca2+সাথে যোগাযোগ ট্রপোনিন, যা এর গঠন এবং ট্রপোমায়োসিনের স্থানচ্যুতির দিকে নিয়ে যায়, যার ফলে অ্যাক্টিন ফিলামেন্ট হয় সক্রিয় এলাকা উন্মুক্ত করা হয়যার সাথে তারা সংযুক্ত মায়োসিন সেতুর প্রধান।
4. Actin সঙ্গে মিথস্ক্রিয়া ফলে মায়োসিন ফিলামেন্টের মাথার ATPase কার্যকলাপ উন্নত করা হয়, এটিপি শক্তি মুক্তি প্রদান, যা ব্যয় করা হয় মায়োসিন সেতুর বাঁকবাহ্যিকভাবে রোয়িং (রোয়িং মুভমেন্ট) (চিত্র 23 দেখুন) এর নড়াচড়ার সাথে সাদৃশ্যপূর্ণ মায়োসিন ফিলামেন্টের তুলনায় অ্যাক্টিন ফিলামেন্টের স্লাইডিং প্রদান করে. একটি স্ট্রোক সম্পূর্ণ করতে এটি একটি ATP অণুর শক্তি লাগে। এই ক্ষেত্রে, সংকোচনশীল প্রোটিনের স্ট্র্যান্ডগুলি 20 এনএম দ্বারা স্থানচ্যুত হয়। মায়োসিন মাথার অন্য অংশে একটি নতুন ATP অণুর সংযুক্তি তার বাগদানের সমাপ্তির দিকে নিয়ে যায়, কিন্তু ATP-এর শক্তি খরচ হয় না। এটিপির অনুপস্থিতিতে, মায়োসিনের মাথাগুলি অ্যাক্টিন থেকে দূরে সরে যেতে পারে না - পেশী টান হয়; যেমন, বিশেষ করে, কঠোর মরটিসের প্রক্রিয়া।
5. এর পর ক্রস ব্রিজগুলির মাথা, তাদের স্থিতিস্থাপকতার কারণে, তাদের আসল অবস্থানে ফিরে আসে এবং পরবর্তী অ্যাক্টিন সাইটের সাথে যোগাযোগ স্থাপন করে; তারপরে অ্যাক্টিন এবং মায়োসিন ফিলামেন্টের আরেকটি রোয়িং আন্দোলন এবং স্লাইডিং আবার ঘটে। এই ধরনের প্রাথমিক কাজগুলি বহুবার পুনরাবৃত্তি হয়। একটি স্ট্রোক (এক ধাপ) প্রতিটি সারকোমেরের দৈর্ঘ্য 1% হ্রাস করে। 50% লোড ছাড়াই একটি বিচ্ছিন্ন ব্যাঙের পেশীর সংকোচনের সাথে, সারকোমেরেসের সংক্ষিপ্তকরণ 0.1 সেকেন্ডে ঘটে। এর জন্য 50টি রোয়িং আন্দোলনের প্রয়োজন।
পেশী সংকোচনের প্রক্রিয়া
মায়োসিন ব্রিজগুলি অ্যাসিঙ্ক্রোনাসভাবে বাঁকে, কিন্তু তাদের মধ্যে অনেকগুলি থাকার কারণে এবং প্রতিটি মায়োসিন ফিলামেন্ট বেশ কয়েকটি অ্যাক্টিন ফিলামেন্ট দ্বারা বেষ্টিত থাকার কারণে, পেশী সংকোচন মসৃণভাবে ঘটে।
শিথিলতাপেশী বিপরীত ক্রমে ঘটমান প্রক্রিয়ার কারণে হয়. সারকোলেমা এবং টি-টিউবিউলের পুনঃপোলারাইজেশনের ফলে এসপিআর ঝিল্লির ক্যালসিয়াম ভোল্টেজ-গেটেড চ্যানেলগুলি বন্ধ হয়ে যায়। Ca-পাম্পগুলি Ca2+ কে SPR-এ ফেরত দেয় (মুক্ত আয়নগুলির ঘনত্ব বৃদ্ধির সাথে পাম্পগুলির কার্যকলাপ বৃদ্ধি পায়)।
ইন্টারফাইব্রিলার স্পেসে Ca2+ ঘনত্বের হ্রাস ট্রপোনিনের বিপরীত রূপের সৃষ্টি করে, যার ফলস্বরূপ ট্রপোমায়োসিন ফিলামেন্টগুলি অ্যাক্টিন ফিলামেন্টের সক্রিয় স্থানগুলিকে বিচ্ছিন্ন করে, যা মায়োসিন ক্রস-ব্রিজের প্রধানগুলির পক্ষে তাদের সাথে যোগাযোগ করা অসম্ভব করে তোলে। বিপরীত দিকে মায়োসিন ফিলামেন্টের সাথে অ্যাক্টিন ফিলামেন্টের স্লাইডিং মাধ্যাকর্ষণ শক্তি এবং পেশী ফাইবার উপাদানগুলির স্থিতিস্থাপক ট্র্যাকশনের ক্রিয়ায় ঘটে, যা সারকোমেরেসের মূল মাত্রা পুনরুদ্ধার করে।
এটিপি কঙ্কালের পেশীগুলির কাজ নিশ্চিত করার জন্য শক্তির উত্স, যার খরচগুলি উল্লেখযোগ্য। এমনকি পেশীগুলির কার্যকারিতার জন্য প্রধান বিনিময়ের শর্তেও, শরীর তার সমস্ত শক্তি সংস্থানগুলির প্রায় 25% প্রভাবিত করে। শারীরিক পরিশ্রমের সময় শক্তি ব্যয় নাটকীয়ভাবে বৃদ্ধি পায়।
পেশী ফাইবারে ATP এর মজুদ নগণ্য (5 mmol/l) এবং 10 টির বেশি একক সংকোচন প্রদান করতে পারে না।
শক্তি খরচনিম্নলিখিত প্রক্রিয়াগুলির জন্য ATP প্রয়োজন।
প্রথমত, ATP-এর শক্তি ব্যয় করা হয় Na/K-পাম্পের ক্রিয়াকলাপ নিশ্চিত করতে (এটি কোষের ভিতরে এবং বাইরে Na+ এবং K+ ঘনত্বের গ্রেডিয়েন্ট বজায় রাখে, যা PP এবং PD গঠন করে, যা ইলেক্ট্রোমেকানিক্যাল কাপলিং প্রদান করে) এবং এর অপারেশন Ca-পাম্প, যা পেশী ফাইবার সংকোচনের পরে সারকোপ্লাজমে Ca2+ ঘনত্ব কমিয়ে দেয়, যা শিথিলতার দিকে পরিচালিত করে।
দ্বিতীয়ত, এটিপির শক্তি মায়োসিন সেতুর (তাদের নমন) রোয়িং আন্দোলনে ব্যয় করা হয়।
এটিপি পুনঃসংশ্লেষণশরীরের তিনটি শক্তি সিস্টেমের সাহায্যে বাহিত.
1. ফসফোজেনিক এনার্জি সিস্টেম পেশীতে উপস্থিত অত্যন্ত শক্তি-নিবিড় সিপি এবং ক্রিয়েটাইন (কে) গঠনের সাথে এটিপি ভেঙে যাওয়ার সময় গঠিত অ্যাডেনোসিন ডিফসফেট (এডিনোসিন ডিফসফেট, এডিপি) এর কারণে এটিপি-র পুনঃসংশ্লেষণ নিশ্চিত করে: ADP + + CF → ATP + K। এটি তাত্ক্ষণিক এটিপি পুনর্সংশ্লেষণ, যখন পেশী উচ্চ শক্তি বিকাশ করতে পারে, তবে অল্প সময়ের জন্য - 6 সেকেন্ড পর্যন্ত, যেহেতু পেশীতে সিএফের মজুদ সীমিত।
2. অ্যানেরোবিক গ্লাইকোলাইটিক এনার্জি সিস্টেম অ্যানারোবিক গ্লুকোজ থেকে ল্যাকটিক অ্যাসিডের ভাঙ্গনের শক্তির কারণে এটিপি রিসিন্থেসিস প্রদান করে। এটিপি পুনঃসংশ্লেষণের এই পথটি দ্রুত, তবে স্বল্পস্থায়ী (1-2 মিনিট), যেহেতু ল্যাকটিক অ্যাসিড জমা হওয়া গ্লাইকোলাইটিক এনজাইমের কার্যকলাপকে বাধা দেয়। যাইহোক, ল্যাকটেট, একটি স্থানীয় ভাসোডিলেটিং প্রভাব সৃষ্টি করে, কার্যকারী পেশীতে রক্ত প্রবাহ উন্নত করে এবং এতে অক্সিজেন এবং পুষ্টি সরবরাহ করে।
3. অ্যারোবিক এনার্জি সিস্টেমের সাহায্যে এটিপি রিসিন্থেসিস প্রদান করে কার্বোহাইড্রেট এবং ফ্যাটি অ্যাসিডের অক্সিডেটিভ ফসফোরিলেশনপেশী কোষের মাইটোকন্ড্রিয়ায় ঘটে। এই পথে কয়েক ঘন্টার জন্য পেশী কাজের জন্য শক্তি প্রদান করতে পারেএবং কঙ্কালের পেশীগুলির কাজের জন্য শক্তি সরবরাহ করার প্রধান উপায়।
পেশী সংকোচনের প্রকার
সংকোচনের প্রকৃতির উপর নির্ভর করেপেশী তিন ধরনের আছে: আইসোমেট্রিক, আইসোটোনিক এবং অক্সোটোনিক.
পেশীর অক্সোটোনিক সংকোচন পেশীর দৈর্ঘ্য এবং টান একযোগে পরিবর্তনের মধ্যে থাকে। এই ধরনের সংকোচন প্রাকৃতিক মোটর ক্রিয়াকলাপের জন্য সাধারণ এবং এটি দুই ধরনের হতে পারে: উদ্ভট, যখন পেশীর টান লম্বা হওয়ার সাথে সাথে থাকে - উদাহরণস্বরূপ, স্কোয়াটিং (নিচু করার) প্রক্রিয়ায় এবং ঘনকেন্দ্রিক, যখন পেশীর টান তার সাথে থাকে। সংক্ষিপ্তকরণ - উদাহরণস্বরূপ, স্কোয়াট করার পরে নীচের অঙ্গগুলি প্রসারিত করার সময় ( আরোহণ)।
আইসোমেট্রিক পেশী সংকোচন- যখন পেশীর টান বৃদ্ধি পায় এবং এর দৈর্ঘ্য পরিবর্তন হয় না। পরীক্ষায় এই ধরণের সংকোচন লক্ষ্য করা যায়, যখন পেশীর উভয় প্রান্ত স্থির থাকে এবং তাদের একত্রিত হওয়ার কোন সম্ভাবনা থাকে না, এবং প্রাকৃতিক পরিস্থিতিতে - উদাহরণস্বরূপ, স্কোয়াটিং এবং অবস্থান ঠিক করার প্রক্রিয়ায়।
আইসোটোনিক পেশী সংকোচনএটা তার ধ্রুবক টান সঙ্গে পেশী ছোট করা গঠিত. এই ধরনের সংকোচন ঘটে যখন একটি সংযুক্ত টেন্ডন সহ একটি আনলোড করা পেশী কোনো বাহ্যিক ভার উত্তোলন (চলানো) ছাড়াই সংকুচিত হয় বা ত্বরণ ছাড়াই একটি লোড উত্তোলন করে।
সময়কালের উপর নির্ভর করেপেশী সংকোচন দুই ধরনের আছে: নির্জন এবং টিটানিক।
একক পেশী সংকোচনস্নায়ু বা পেশী নিজেই একটি একক জ্বালা সঙ্গে ঘটে. সাধারণত পেশী তার মূল দৈর্ঘ্যের 5-10% দ্বারা সংক্ষিপ্ত হয়। একক সংকোচন বক্ররেখায় তিনটি প্রধান পর্যায় রয়েছে: 1) সুপ্ত- জ্বালা প্রয়োগের মুহূর্ত থেকে সংকোচনের শুরু পর্যন্ত সময়; 2) সময়কাল সংক্ষিপ্তকরণ (বা চাপের বিকাশ); 3) সময়কাল শিথিলকরণ. একক মানুষের পেশী সংকোচনের সময়কাল পরিবর্তনশীল। উদাহরণস্বরূপ, সোলিয়াস পেশীতে, এটি 0.1 সেকেন্ড। সুপ্ত সময়ের মধ্যে, পেশী তন্তুগুলির উত্তেজনা এবং ঝিল্লি বরাবর এটির সঞ্চালন হয়। পেশী ফাইবারের একক সংকোচনের সময়কালের অনুপাত, এর উত্তেজনা এবং পেশী ফাইবারের উত্তেজনায় ফেজ পরিবর্তনগুলি চিত্রে দেখানো হয়েছে। 24.
পেশী ফাইবার সংকোচনের সময়কাল AP-এর তুলনায় অনেক বেশি, কারণ Ca-পাম্পগুলি Ca2+ কে SPR এবং পরিবেশে ফিরিয়ে আনতে কাজ করতে সময় নেয় এবং ইলেক্ট্রোফিজিওলজিকালের তুলনায় যান্ত্রিক প্রক্রিয়াগুলির বৃহত্তর জড়তা।

ভাত। চিত্র 24. উষ্ণ-রক্তযুক্ত কঙ্কালের পেশীর ধীর ফাইবারের AP (A) এবং একটি একক সংকোচন (B) হওয়ার সময়ের অনুপাত। তীর- বিরক্তির মুহূর্ত। দ্রুত তন্তুগুলির সংকোচনের সময় কয়েকগুণ কম
টিটানিক সংকোচন- এটি একটি দীর্ঘায়িত পেশী সংকোচন যা ছন্দবদ্ধ উদ্দীপনার প্রভাবে ঘটে, যখন প্রতিটি পরবর্তী জ্বালা বা স্নায়ু আবেগ পেশীতে আসে যখন এটি এখনও শিথিল হয়নি। টেটানিক সংকোচন একক পেশী সংকোচনের সংমিশ্রণের ঘটনার উপর ভিত্তি করে (চিত্র 25) - একটি পেশী ফাইবার বা দুটি বা ততোধিক দ্রুত পেশীর সম্পূর্ণ পেশীতে প্রয়োগ করা হলে সংকোচনের প্রশস্ততা এবং সময়কাল বৃদ্ধি পায়।

ভাত। চিত্র 25. ব্যাঙের গ্যাস্ট্রোকনেমিয়াস পেশীর সংকোচনের সমষ্টি: 1 - শিথিল পেশীর প্রথম জ্বালার প্রতিক্রিয়ায় একক সংকোচন বক্ররেখা; 2 - দ্বিতীয় উদ্দীপনার প্রতিক্রিয়ায় একই পেশীর একতরফা সংকোচনের বক্ররেখা; 3 - সংকোচনকারী পেশীর জোড়া উদ্দীপনার ফলে প্রাপ্ত সমষ্টি সংকোচনের বক্ররেখা ( তীর দ্বারা নির্দেশিত)
এই ক্ষেত্রে, পূর্ববর্তী সংকোচনের সময়কালে জ্বালা হওয়া উচিত। সংকোচনের প্রশস্ততা বৃদ্ধি পেশী তন্তুগুলির বারবার উত্তেজনার সময় হায়ালোপ্লাজমে Ca2+ এর ঘনত্বের বৃদ্ধি দ্বারা ব্যাখ্যা করা হয়, যেহেতু Ca-পাম্পের এটি SPR-এ ফেরত দেওয়ার সময় নেই। Ca2+ অ্যাক্টিন ফিলামেন্টের সাথে মায়োসিন ব্রিজের ব্যস্ততার জোনের সংখ্যা বৃদ্ধি করে।
যদি বারবার আবেগ বা বিরক্তিগুলি পেশী শিথিলতার পর্যায়ে প্রবেশ করে, সেখানে আছে দাঁতের টিটেনাস. যদি বারবার irritations সংক্ষিপ্ত পর্যায়ে পড়া, আছে মসৃণ টিটেনাস(চিত্র 26)।

ভাত। 26. সায়াটিক স্নায়ুর উদ্দীপনার বিভিন্ন ফ্রিকোয়েন্সিতে ব্যাঙের গ্যাস্ট্রোকনেমিয়াস পেশীর সংকোচন: 1 - একক সংকোচন (ফ্রিকোয়েন্সি 1 Hz); 2,3 - দানাদার টিটেনাস (15-20 Hz); 4.5 - মসৃণ টিটেনাস (25-60 Hz); 6 - উদ্দীপনার অস্বস্তিকর ফ্রিকোয়েন্সিতে শিথিলতা (120 Hz)
সংকোচনের প্রশস্ততা এবং মসৃণ টিটেনাস সহ পেশী তন্তুগুলির দ্বারা বিকশিত উত্তেজনার পরিমাণ সাধারণত একক সংকোচনের চেয়ে 2-4 গুণ বেশি হয়। পেশী তন্তুগুলির টিটানিক সংকোচন, একক সংকোচনের বিপরীতে, তাদের ক্লান্তি দ্রুত ঘটায়।
স্নায়ু বা পেশীর উদ্দীপনার ফ্রিকোয়েন্সি বৃদ্ধির সাথে, মসৃণ টিটেনাসের প্রশস্ততা বৃদ্ধি পায়। সর্বোচ্চ টিটেনাস নামকরণ করা হয় সর্বোত্তমটিটেনাসের বৃদ্ধি হায়ালোপ্লাজমে Ca2+ জমা হওয়ার দ্বারা ব্যাখ্যা করা হয়। স্নায়ু উদ্দীপনার ফ্রিকোয়েন্সি আরও বৃদ্ধির সাথে (প্রায় 100 Hz), নিউরোমাসকুলার সিন্যাপসে উত্তেজনা সঞ্চালনে একটি ব্লকের বিকাশের কারণে পেশী শিথিল হয় - ভেদেনস্কির হতাশা(জ্বালা ফ্রিকোয়েন্সি হতাশাবাদী) (চিত্র 26 দেখুন)। Vvedensky pessimum এছাড়াও পেশীর প্রত্যক্ষ, কিন্তু আরো ঘন ঘন উদ্দীপনা (প্রায় 200 ডাল/সে) দ্বারা প্রাপ্ত করা যেতে পারে, যাইহোক, পরীক্ষার বিশুদ্ধতার জন্য, নিউরোমাসকুলার সিন্যাপ্সগুলিকে অবরুদ্ধ করা উচিত। যদি, পেসিমাম হওয়ার পরে, উদ্দীপনার ফ্রিকোয়েন্সি সর্বোত্তমটিতে হ্রাস করা হয়, তবে পেশী সংকোচনের প্রশস্ততা তাত্ক্ষণিকভাবে বৃদ্ধি পায় - প্রমাণ যে পেসিমাম পেশী ক্লান্তি বা শক্তি সংস্থান হ্রাসের ফলাফল নয়।
প্রাকৃতিক অবস্থার অধীনে, পৃথক পেশী তন্তুগুলি প্রায়শই দানাদার টিটেনাস মোডে সংকোচন করে, তবে, তাদের সংকোচনের অসিঙ্ক্রোনির কারণে পুরো পেশীর সংকোচন একটি মসৃণ টিটেনাসের মতো হয়।
ফেডারেল এজেন্সি ফর এডুকেশন
পেনজা স্টেট পেডাগজিকাল ইউনিভার্সিটি V.G এর পরে নামকরণ করা হয়েছে। বেলিন্সকি
প্রাকৃতিক ভূগোল প্রোটোকল নং ___ তারিখ "___" _________ 2006 অনুষদের একাডেমিক কাউন্সিলের সভায় গৃহীত
অনুষদের ডিন _______________
এল.ভি. ক্রিভোশেভা আমি অনুমোদন করি
একাডেমিক বিষয়ক ভাইস রেক্টর
______________________________
এম.এ. পিয়াটিন
শিক্ষামূলক কাজের প্রোগ্রাম
শৃঙ্খলায় "পেশী সংকোচনের জৈব রসায়ন"
বিশেষত্বের জন্য
020208 (012300) - "বায়োকেমিস্ট্রি"
ন্যাচারাল জিওগ্রাফিক অনুষদ
বায়োকেমিস্ট্রি বিভাগ
পেনজা, 2006
শৃঙ্খলার জন্য রাষ্ট্রীয় শিক্ষাগত মানদণ্ডের প্রয়োজনীয়তা
সূচক
যোগ্যতার প্রয়োজনীয়তা
একজন বিশেষজ্ঞ বায়োকেমিস্টের প্রস্তুতি জৈবিক অনুষদ বা বিভাগগুলিতে বায়োকেমিস্ট্রি বিভাগে বাহিত হয়। একজন বায়োকেমিস্ট বিশেষজ্ঞের প্রধান শিক্ষামূলক কর্মসূচির বাস্তবায়ন শিক্ষকদের দ্বারা প্রাথমিক শিক্ষা এবং/অথবা কাজের অভিজ্ঞতা এবং প্রকাশনাগুলি দ্বারা নিশ্চিত করা বৈজ্ঞানিক এবং বৈজ্ঞানিক-পদ্ধতিগত কাজ পদ্ধতিগতভাবে পরিচালনা করা, শেখানো শাখাগুলির প্রোফাইলে প্রদান করা উচিত। একাডেমিক ডিগ্রি এবং শিরোনাম সহ শিক্ষকদের অনুপাত কমপক্ষে 67% হওয়া উচিত। বিশেষ শাখার শিক্ষকদের, একটি নিয়ম হিসাবে, প্রাসঙ্গিক পেশাগত ক্ষেত্রে একটি একাডেমিক ডিগ্রি এবং অভিজ্ঞতা থাকতে হবে।
শৃঙ্খলার লক্ষ্য এবং উদ্দেশ্য
"পেশী সংকোচনের বায়োকেমিস্ট্রি" কোর্সটি শিক্ষার্থীদের আণবিক গঠন এবং স্বাভাবিক এবং প্যাথলজিকাল পরিস্থিতিতে পেশীগুলির কার্যকারিতার মূল বিষয়গুলির সাথে সাথে শারীরিক অনুশীলন এবং খেলাধুলার সময় তাদের মধ্যে ঘটে যাওয়া জৈবিক প্রক্রিয়াগুলির বৈশিষ্ট্যগুলির সাথে পরিচিত করা উচিত।
পেশী সংকোচনের বায়োকেমিস্ট্রি হল জৈব রসায়নের একটি শাখা এবং এটি তাত্ত্বিক এবং পরীক্ষামূলক পদার্থবিদ্যা, আণবিক জীববিজ্ঞান, মানব দেহবিজ্ঞান এবং চিকিৎসা শাখার সংযোগস্থলে অবস্থিত।
কোর্সের বিষয়বস্তুতে পেশী টিস্যুতে প্রোটিনের গঠন এবং ক্রিয়াকলাপের বৈশিষ্ট্য, পেশী কার্যকলাপের শক্তি সরবরাহ এবং স্বাভাবিক অবস্থায় জৈব রাসায়নিক প্রক্রিয়াগুলির পাশাপাশি ব্যায়াম এবং প্যাথলজির সময় বিষয়গুলি রয়েছে। এই কোর্সের অধ্যয়ন ভবিষ্যতের বিশেষজ্ঞদের আণবিক স্তরে শরীরের পেশী টিস্যুতে ঘটে যাওয়া প্রক্রিয়াগুলির সারমর্ম এবং তাত্পর্য বুঝতে দেয়।
কোর্সের উদ্দেশ্য: মানব জৈব রসায়নের মৌলিক তথ্য এবং শারীরিক শিক্ষার অনুশীলনে এই জ্ঞান ব্যবহারের সম্ভাবনা একত্রিত করা।
কোর্সের উদ্দেশ্য:
পেশী কার্যকলাপের সময় বিপাকের বৈশিষ্ট্য সম্পর্কে আধুনিক তথ্য উপস্থাপন;
জৈব রাসায়নিক ভিত্তি প্রকাশ করুন:
ব্যায়ামের সময় শরীরের পরিবর্তন
ক্লান্তি প্রক্রিয়া
পুনরুদ্ধার প্রক্রিয়া
প্রশিক্ষণ অভিযোজন
প্রশিক্ষণ পদ্ধতি
প্রশিক্ষণ প্রক্রিয়ার কার্যকারিতা
ক্রীড়া কর্মক্ষমতা
মোটর গুণাবলী এবং ক্রীড়াবিদদের সহনশীলতার বিকাশ
পুষ্টি
ক্রীড়াবিদদের অবস্থা পর্যবেক্ষণ
প্রোগ্রামটি স্পেশালিটি 020208 (012300) "বায়োকেমিস্ট্রি" এ নথিভুক্ত শিক্ষার্থীদের জন্য উচ্চতর পেশাগত শিক্ষার রাজ্য শিক্ষাগত মান অনুযায়ী সংকলিত হয়েছে।
এই বিশেষত্বের পাঠ্যক্রম অনুসারে, "পেশী সংকোচনের বায়োকেমিস্ট্রি" কোর্সের জন্য 68 ঘন্টা বরাদ্দ করা হয়, যার মধ্যে 34 ঘন্টা শ্রেণীকক্ষের জন্য এবং 34 ঘন্টা স্বাধীন কাজের জন্য। ক্লাসরুমের 34 ঘন্টা কাজের মধ্যে 34 ঘন্টাই বক্তৃতা। কোর্সটি একটি ক্রেডিট প্রদান করে।
সেমিস্টার এবং শিক্ষার ধরন অনুসারে শেখার সময় বন্টন
সেমিস্টার
শৃঙ্খলা বিষয়বস্তু
ভূমিকা
পেশী সংকোচনের জৈব রসায়ন বিষয়। কোর্সের উদ্দেশ্য এবং বিষয়বস্তু। সংক্ষিপ্ত ঐতিহাসিক ওভারভিউ। বায়োকেমিস্টদের প্রশিক্ষণের বিষয় হিসাবে পেশী সংকোচনের জৈব রসায়নের মূল্য। প্রাকৃতিক বিজ্ঞানের সাধারণ ব্যবস্থায় পেশী সংকোচনের জৈব রসায়নের অবস্থান।
সাইটোপ্লাজমের পেশীবহুল সিস্টেম
সাইটোস্কেলটনের সংকোচনশীল প্রোটিন।
তন্তুযুক্ত কাঠামোর ধরন: মাইক্রোফিলামেন্ট, মাইক্রোটিউবুলস, মধ্যবর্তী ফিলামেন্ট। তাদের গঠন এবং ফাংশন নিয়ন্ত্রণ.
সিলিয়ার গঠন এবং চলাচল। ব্যাকটেরিয়ার মোটর যন্ত্রপাতি।
কোলাজেন
কোলাজেনের প্রকার। কোলাজেন অণুর গঠনের বৈশিষ্ট্য। কোলাজেনের সংশ্লেষণ। কোলাজেন অস্বাভাবিকতার কারণে বংশগত রোগ।
পেশী টিস্যুর গঠন এবং রাসায়নিক গঠন
পেশী টিস্যুর শ্রেণীবিভাগ।
স্ট্রাইটেড পেশীর আকারগত সংগঠন। পেশী ফাইবারের গঠন। স্ট্রাইটেড পেশীর রাসায়নিক গঠন। পেশী প্রোটিন যা সারকোপ্লাজম তৈরি করে: মায়োগ্লোবিন, পারভালবুমিন। সংকোচনশীল প্রোটিন: মায়োসিন, অ্যাক্টিন, অ্যাক্টোমায়োসিন, ট্রপোমায়োসিন, ট্রপোনিন টি, ট্রপোনিনআমি, ট্রপোনিন সি, - এবং -অ্যাক্টিন। অ-প্রোটিন নাইট্রোজেনাস নিষ্কাশন। নাইট্রোজেন মুক্ত পদার্থ।
হৃৎপিণ্ডের পেশী এবং মসৃণ পেশীগুলির রাসায়নিক গঠনের বৈশিষ্ট্য।
অনটোজেনেসিসে পেশী টিস্যুর রাসায়নিক গঠনের পরিবর্তন।
5. কার্যকরী পেশী বায়োকেমিস্ট্রি
পেশী সংকোচনের জৈব রাসায়নিক চক্র। পেশী সংকোচন এবং শিথিলকরণের নিয়ন্ত্রণ: স্ট্রাইটেড পেশীগুলির অ্যাক্টিন নিয়ন্ত্রণ, মসৃণ পেশীগুলির মায়োসিন নিয়ন্ত্রণ। ক্যালসিয়াম আয়ন এবং মডুলেটরি প্রোটিনের ভূমিকা।
6. পেশীবহুল কার্যকলাপের সময় বায়োএনার্জেটিক প্রক্রিয়া
পেশী ক্রিয়াকলাপের সময় অ্যানেরোবিক (মায়োকিনেজ, ক্রিয়েটাইন ফসফোকিনেস এবং গ্লাইকোলাইটিক) এবং অ্যারোবিক (অক্সিডেটিভ ফসফোরিলেশন) এটিপি পুনঃসংশ্লেষণের পথ। বিভিন্ন শক্তি এবং তীব্রতার পেশী কার্যকলাপের সময় এটিপি পুনঃসংশ্লেষণের বিভিন্ন পথের অনুপাত। বিভিন্ন শক্তির উত্স চালু করার ক্রম।
7. ব্যায়াম এবং খেলাধুলার বায়োকেমিস্ট্রি
অক্সিজেন পরিবহনের জৈব রাসায়নিক বৈশিষ্ট্য এবং পেশীতে এর জমা। কর্মক্ষেত্রে অক্সিজেন খরচ, অক্সিজেনের ঘাটতি এবং অক্সিজেন ঋণ। শক্তি খরচ এবং ব্যায়াম অক্সিজেন চাহিদা.
জৈব রাসায়নিক পরিবর্তন যা প্রশিক্ষণের প্রভাবে পেশীতন্ত্র, অভ্যন্তরীণ অঙ্গ, রক্ত এবং কেন্দ্রীয় স্নায়ুতন্ত্রে ঘটে। স্ট্যান্ডার্ড এবং সর্বাধিক কাজের সময় জৈব রাসায়নিক পরিবর্তন, ক্লান্তি সহ, বিশ্রামের সময়কালে। প্রশিক্ষণ এবং detraining সময় জৈব রাসায়নিক পরিবর্তনের ক্রম. অতিরিক্ত প্রশিক্ষণের সময় জৈব রাসায়নিক পরিবর্তন।
আপেক্ষিক শক্তি অনুযায়ী ক্রীড়া অনুশীলনের শ্রেণীবিভাগের জৈব রাসায়নিক প্রমাণ। সাইক্লিক (দৌড়ানো, সাঁতার, সাইক্লিং, স্কিইং এবং স্কেটিং) এবং অ্যাসাইক্লিক (ভারোত্তোলন, বক্সিং, জিমন্যাস্টিকস, কুস্তি) ব্যায়ামের সময় শরীরে জৈব রাসায়নিক পরিবর্তন।
8. প্যাথলজিতে পেশীতে জৈব রাসায়নিক পরিবর্তন
বিষয় অনুসারে ঘন্টার আনুমানিক বন্টন
p/n
কন্ট্রোল কাজের উদাহরণ বিষয়
1. পেশী টিস্যুর প্রকারভেদ। পেশী গঠন। পেশী সংকোচনের বায়োকেমিস্ট্রি। পেশী কার্যকলাপের সময় বিপাক - পরীক্ষা।
সাহিত্য
প্রধান:
অতিরিক্ত:
2. বিভিন্ন শক্তি এবং সময়কালের ব্যায়াম করার সময় শরীরে জৈব রাসায়নিক পরিবর্তন। ক্লান্তি এবং পুনরুদ্ধারের জৈব রাসায়নিক কারণ। ক্রীড়া প্রশিক্ষণের সময় জৈব রাসায়নিক অভিযোজনের নিদর্শন। সহনশীলতার জৈব রাসায়নিক ভিত্তি, গতি-শক্তি গুণাবলী, ক্রীড়া কর্মক্ষমতা। - সমস্যা সমাধান.
সাহিত্য
প্রধান:
1. অ্যালবার্টস বি., ব্রে ডি., লুইস জে., রাফ এম., রবার্টস কে., ওয়াটসন জে. কোষের আণবিক জীববিজ্ঞান৷ 3 খণ্ডে। ২য় সংস্করণ। - এম.: মীর, 1994
2. বেরেজভ T.T., Korovkin B.F. জৈবিক রসায়ন। - এম.: মেডিসিন, 2002
3. মারি আর., গ্রেনার ডি., মেইস পি., রডওয়েল ভি. হিউম্যান বায়োকেমিস্ট্রি: পার। ইংরেজী থেকে. - এম.: মীর, 1993
4. বায়োকেমিস্ট্রি / এড. মেনশিকোভা V.V., Volkova N.I. - এম।: শারীরিক সংস্কৃতি এবং খেলাধুলা, 1986
5. Volkov N.I. পেশী কার্যকলাপের বায়োকেমিস্ট্রি। - এম.: অলিম্পিক খেলা, 2001।
6. ইয়াকভলেভ এন.এন. খেলাধুলার বায়োকেমিস্ট্রি। - এম।: শারীরিক সংস্কৃতি এবং খেলাধুলা, 1974
7. ইয়াকভলেভ এন.এন. গতির রসায়ন। - এল.: নাউকা, 1983
অতিরিক্ত:
1. হিউম্যান ফিজিওলজি / কোসিটস্কি জি.আই. – এম.: মেডিসিন, 1985, 544 পি।
2. বায়োকেমিস্ট্রি এবং মলিকুলার বায়োলজি / এলিয়ট ভি., এলিয়ট ডি.; প্রতি ইংরেজি থেকে: O.V. ডব্রিনিনা এবং অন্যান্য; এড. A.I. আর্চাকোভা - এম।: MAIS "নাউকা / ইন্টারপেরিওডিকা", 2002, 446s।
3. লেহনিঙ্গার এ. বায়োকেমিস্ট্রি। টি. 1 - 3. এম.: মীর, 1985
জ্ঞানের চূড়ান্ত নিয়ন্ত্রণের ফর্ম - পরীক্ষা
অফসেটের জন্য প্রশ্নের উদাহরণের তালিকা
পেশী কার্যকলাপের সময় বিপাকের বৈশিষ্ট্য।
পেশী এবং পেশী তন্তুগুলির প্রকার। পেশী তন্তুগুলির কাঠামোগত সংগঠন। পেশী টিস্যুর রাসায়নিক গঠন।
সংকোচন এবং শিথিলকরণের সময় পেশীগুলির কাঠামোগত এবং জৈব রাসায়নিক পরিবর্তন। পেশী সংকোচনের আণবিক প্রক্রিয়া।
পেশী সংকোচন এবং শিথিলকরণের নিয়ন্ত্রণ: স্ট্রাইটেড পেশীগুলির অ্যাক্টিন নিয়ন্ত্রণ, মসৃণ পেশীগুলির মায়োসিন নিয়ন্ত্রণ। ক্যালসিয়াম আয়ন এবং মডুলেটরি প্রোটিনের ভূমিকা।
শক্তি উৎপাদন প্রক্রিয়ার সাধারণ বৈশিষ্ট্য। ক্রিয়েটাইন ফসফোকিনেস, গ্লাইকোলাইটিক, মায়োকিনেস, এটিপি রিসিন্থেসিস এর বায়বীয় প্রক্রিয়া। বিভিন্ন শক্তি এবং তীব্রতার পেশী কার্যকলাপের সময় এটিপি পুনঃসংশ্লেষণের বিভিন্ন পথের অনুপাত। বিভিন্ন শক্তির উত্স চালু করার ক্রম।
পেশী কার্যকলাপের সময় জৈব রাসায়নিক প্রক্রিয়ার পরিবর্তনের সাধারণ দিক। কর্মক্ষম পেশীতে অক্সিজেন পরিবহন এবং পেশী কার্যকলাপের সময় এর ব্যবহার।
পেশীবহুল কাজের সময় পৃথক অঙ্গ এবং টিস্যুতে জৈব রাসায়নিক পরিবর্তন। পেশীবহুল কাজের সময় জৈব রাসায়নিক পরিবর্তনের প্রকৃতি অনুসারে শারীরিক ব্যায়ামের শ্রেণীবিভাগ।
ক্লান্তির জৈব রাসায়নিক কারণ।
পুনরুদ্ধার প্রক্রিয়ার জৈব রাসায়নিক ভিত্তি।
যে বিষয়গুলো একজন ব্যক্তির শারীরিক কর্মক্ষমতা সীমিত করে। অ্যাথলিটের অ্যারোবিক এবং অ্যানেরোবিক পারফরম্যান্সের সূচক।
ক্রীড়াবিদদের কর্মক্ষমতা উপর প্রশিক্ষণ প্রভাব. বয়স এবং অ্যাথলেটিক পারফরম্যান্স।
গতি-শক্তি গুণাবলীর জৈব রাসায়নিক বৈশিষ্ট্য। ক্রীড়াবিদদের গতি-শক্তি প্রশিক্ষণের পদ্ধতির জৈব রাসায়নিক ভিত্তি।
সহনশীলতার জৈব রাসায়নিক কারণ। প্রশিক্ষণের পদ্ধতি যা সহনশীলতার বিকাশে অবদান রাখে।
শারীরিক কার্যকলাপ, অভিযোজন এবং প্রশিক্ষণ প্রভাব। জৈব রাসায়নিক অভিযোজন এবং প্রশিক্ষণের নীতিগুলির বিকাশের নিদর্শন। প্রশিক্ষণের সময় শরীরে অভিযোজিত পরিবর্তনের নির্দিষ্টতা।
প্রশিক্ষণের সময় অভিযোজিত পরিবর্তনের বিপরীততা। প্রশিক্ষণের সময় অভিযোজিত পরিবর্তনের ক্রম।
প্রশিক্ষণ সময় প্রশিক্ষণ প্রভাব মিথস্ক্রিয়া. প্রশিক্ষণের প্রক্রিয়ায় অভিযোজনের চক্রাকার বিকাশ।
ক্রীড়াবিদদের যুক্তিসঙ্গত পুষ্টির নীতি। শরীরের শক্তি খরচ এবং সম্পাদিত কাজের উপর তার নির্ভরতা। ক্রীড়াবিদদের খাদ্যে পুষ্টির ভারসাম্য।
পেশী কার্যকলাপ প্রদান খাদ্য পৃথক রাসায়নিক উপাদান ভূমিকা. পুষ্টিকর পরিপূরক এবং শরীরের ওজন নিয়ন্ত্রণ।
জৈব রাসায়নিক নিয়ন্ত্রণের কাজ, প্রকার এবং সংগঠন। অধ্যয়নের বস্তু এবং মৌলিক জৈব রাসায়নিক পরামিতি। রক্ত এবং প্রস্রাবের গঠনের প্রধান জৈব রাসায়নিক সূচক, পেশী কার্যকলাপের সময় তাদের পরিবর্তন। পেশী কার্যকলাপের সময় শরীরের শক্তি সরবরাহ ব্যবস্থার বিকাশের জৈব রাসায়নিক নিয়ন্ত্রণ।
প্রশিক্ষণের স্তরের উপর জৈব রাসায়নিক নিয়ন্ত্রণ, ক্লান্তি এবং ক্রীড়াবিদদের শরীরের পুনরুদ্ধার। খেলাধুলায় ডোপিং নিয়ন্ত্রণ।
২. প্রতিবেদন তৈরি: বিভিন্ন খেলাধুলার জৈব রাসায়নিক ভিত্তি।
পরিকল্পনা:
প্রশিক্ষণের সময় শক্তি সিস্টেমের সংযোগ এবং তাদের অভিযোজন
বায়বীয় এবং অ্যানেরোবিক প্রক্রিয়াগুলির শক্তি এবং ক্ষমতা
শরীরের জৈব রাসায়নিক পরিবর্তন
ক্লান্তি এবং পুনরুদ্ধারের প্রক্রিয়া
প্রশিক্ষণের সময় অভিযোজিত পরিবর্তনের নির্দিষ্টতা
খাদ্য জৈব রসায়ন
শৃঙ্খলার শিক্ষাগত এবং পদ্ধতিগত সহায়তা
অ্যালবার্টস বি., ব্রে ডি., লুইস জে., রাফ এম., রবার্টস কে., ওয়াটসন জে. কোষের আণবিক জীববিজ্ঞান৷ 3 খণ্ডে। ২য় সংস্করণ। - এম.: মীর, 1994
বেরেজভ টি.টি., কোরোভকিন বি.এফ. জৈবিক রসায়ন। - এম.: মেডিসিন, 2002
লেহনিঙ্গার এ. বায়োকেমিস্ট্রি। টি. 1 - 3. এম.: মীর, 1985
মারি আর., গ্রেনার ডি., মেইস পি., রডওয়েল ডব্লিউ. হিউম্যান বায়োকেমিস্ট্রি: পার। ইংরেজী থেকে. - এম.: মীর, 1993
জৈব রসায়ন এবং আণবিক জীববিদ্যা / এলিয়ট ভি., এলিয়ট ডি.; প্রতি ইংরেজি থেকে: O.V. ডব্রিনিনা এবং অন্যান্য; এড. A.I. আর্চাকোভা - এম।: MAIS "নাউকা / ইন্টারপেরিওডিকা", 2002, 446s।
বায়োকেমিস্ট্রি/এড. মেনশিকোভা V.V., Volkova N.I. - এম।: শারীরিক সংস্কৃতি এবং খেলাধুলা, 1986
অতিরিক্ত সাহিত্য
Volkov N.I. পেশী কার্যকলাপের বায়োকেমিস্ট্রি। - এম.: অলিম্পিক খেলা, 2001।
হিউম্যান ফিজিওলজি / কোসিটস্কি জি.আই. – এম.: মেডিসিন, 1985, 544 পি।
ইয়াকভলেভ এন.এন. খেলাধুলার বায়োকেমিস্ট্রি। - এম।: শারীরিক সংস্কৃতি এবং খেলাধুলা, 1974
ইয়াকভলেভ এন.এন. গতির রসায়ন। - এল.: নাউকা, 1983
বিশেষত্ব 0202028 (012300)-এর জন্য "পেশী সংকোচনের বায়োকেমিস্ট্রি" - "বায়োকেমিস্ট্রি" শৃঙ্খলার পাঠ্যক্রমটি বায়োকেমিস্ট্রি বিভাগের একটি সভায় আলোচনা এবং অনুমোদিত হয়েছিল
মিনিট নং _____ তারিখ "____" _____________ 2006
মাথা বায়োকেমিস্ট্রি বিভাগ
জৈবিক বিজ্ঞানের ডক্টর, অধ্যাপক ____________________________________ এম.টি. গেঞ্জিন
(স্বাক্ষর)
প্রাকৃতিক ভূগোল অনুষদের পদ্ধতিগত কাউন্সিল দ্বারা অনুমোদিত
মিনিট নং ____________ তারিখ "_____" ___________ 2006
পদ্ধতিগত কাউন্সিলের চেয়ারম্যান ড
প্রাকৃতিক ভূগোল অনুষদ,
কারিগরি বিজ্ঞানের প্রার্থী, সহযোগী অধ্যাপক _____________________ O.V. জরকিন
(স্বাক্ষর)
দ্বারা কম্পাইল:
ক্যান্ড biol বিজ্ঞান, সহযোগী অধ্যাপক Petrushova O.P. _________________________________
(স্বাক্ষর)
অধ্যায় 3. কার্যকরী পেশী বায়োকেমিস্ট্রি
3.1। পেশী সংকোচনের প্রক্রিয়া
পেশী সিস্টেমের বহুবিধ কার্যকারিতা সত্ত্বেও, পেশীগুলির প্রধান কাজ হল একটি মোটর অ্যাক্টের বাস্তবায়ন, অর্থাৎ সংকোচন এবং শিথিলকরণ। পেশী সংকোচন একটি জটিল যান্ত্রিক রাসায়নিক প্রক্রিয়া যার সময় ATP এর হাইড্রোলাইটিক ভাঙ্গনের রাসায়নিক শক্তি যান্ত্রিক শক্তিতে রূপান্তরিত হয়। আসুন আমরা মেরুদণ্ডী প্রাণীদের স্ট্রাইটেড পেশীগুলির সংকোচনের প্রক্রিয়াটির কাঠামোগত ভিত্তি বিবেচনা করি, যেহেতু এই প্রক্রিয়াটি সবচেয়ে সম্পূর্ণরূপে অধ্যয়ন করা হয়েছে। যেমন উল্লিখিত হয়েছে, স্ট্রাইটেড পেশীর সংকোচন পদ্ধতিতে ওভারল্যাপিং প্রোটিন ফিলামেন্ট থাকে যা একে অপরের সাপেক্ষে স্লাইড করে (চিত্র 9, এ দেখুন)।
ই. হাক্সলে এবং আর. নিডারগারকে, সেইসাথে এক্স. হাক্সলে এবং জে. হেনসনের প্রস্তাবিত মডেল অনুসারে, যখন মায়োফাইব্রিলগুলি সংকুচিত হয়, তখন থ্রেডগুলির একটি সিস্টেম অন্যটিতে প্রবেশ করে, অর্থাৎ, থ্রেডগুলি একে অপরের উপর স্লাইড করতে শুরু করে। এটি ছিল, যা পেশী সংকোচনের কারণ।
ATP এর হাইড্রোলাইসিসের সময় নির্গত শক্তির কারণে সংকোচন ঘটে। একটি স্ট্রাইটেড পেশীতে, সংকোচন Ca 2+ আয়নের ঘনত্বের উপর নির্ভর করে, যা ঘুরেফিরে, সারকোপ্লাজমিক রেটিকুলাম দ্বারা নিয়ন্ত্রিত হয়, একটি বিশেষ ঝিল্লি সিস্টেম যা বিশ্রামে Ca 2+ জমা করে এবং যখন একটি স্নায়ু আবেগ প্রয়োগ করা হয় তখন এটি ছেড়ে দেয়। পেশী ফাইবার (চিত্র 11, A, B দেখুন)।
1) মায়োসিন "হেড" ATP থেকে ADP এবং H 3 PO 4 (P i) কে হাইড্রোলাইজ করতে পারে, কিন্তু হাইড্রোলাইসিস পণ্যের মুক্তি নিশ্চিত করে না। অতএব, এই প্রক্রিয়াটি অনুঘটক প্রকৃতির চেয়ে বরং স্টোইচিওমেট্রিক (চিত্র 10, ক দেখুন);
3) এই মিথস্ক্রিয়া অ্যাক্টিন-মায়োসিন কমপ্লেক্স থেকে ADP এবং H 3 RO 4 এর মুক্তি নিশ্চিত করে। অ্যাক্টোমায়োসিন বন্ডের 45° কোণে সর্বনিম্ন শক্তি থাকে; অতএব, ফাইব্রিল অক্ষের সাথে মায়োসিনের কোণ 90° থেকে 45° (প্রায়) পরিবর্তিত হয় এবং অ্যাক্টিন অগ্রসর হয় (10-15 nm দ্বারা) সারকোমের কেন্দ্রের দিকে (দেখুন চিত্র 10, গ) (চিত্র 9 বি-তে রোয়িং বোটের মডেল অনুসারে "স্ট্রোক");
ভাত। 9. মেরুদণ্ডী প্রাণীদের কঙ্কালের পেশীগুলির সংগঠন
এবং পেশী সংকোচনের প্রক্রিয়া
4) একটি নতুন ATP অণু মায়োসিন-এফ-অ্যাক্টিন কমপ্লেক্সের সাথে আবদ্ধ হয় (চিত্র 10, ডি দেখুন);
5) মায়োসিন-এটিপি কমপ্লেক্সের অ্যাক্টিনের সাথে কম সখ্যতা রয়েছে এবং তাই এফ-অ্যাক্টিন থেকে মায়োসিন (এটিপি) "হেড" এর বিচ্ছেদ ঘটে। শেষ পর্যায়টি আসলে শিথিলকরণ, যা স্পষ্টভাবে অ্যাক্টিন-মায়োসিন কমপ্লেক্সের সাথে ATP-এর আবদ্ধতার উপর নির্ভর করে (চিত্র 10e দেখুন)। তারপর চক্র আবার শুরু হয়।
ভাত। দশপেশী সংকোচনের জৈব রাসায়নিক চক্র
যতক্ষণ এটিপি থাকে ততক্ষণ চক্রটি পুনরাবৃত্তি হয়। 500 পুরু ফিলামেন্ট মায়োসিন "হেডস" এর প্রতিটি "স্ট্রোক" 10 এনএম এর পরিবর্তন ঘটায়। শক্তিশালী সংকোচনের সময়, স্ট্রোকের হার প্রতি সেকেন্ডে প্রায় 5 বার হয়। এটিপি হাইড্রোলাইসিসের প্রতিটি চক্রের সাথে, মায়োসিনের "মাথা" নতুন অ্যাক্টিন অণুর সাথে যোগাযোগ করে, যার কারণে মায়োসিন এবং অ্যাক্টিন ফিলামেন্টগুলির পারস্পরিক স্লাইডিং ঘটে, অর্থাৎ পেশী ফাইবার সংকোচন ঘটে।
3.2। পেশী সংকোচন এবং শিথিলকরণ নিয়ন্ত্রণ
যেকোন পেশীর সংকোচন পূর্বে বর্ণিত সাধারণ প্রক্রিয়া অনুসারে ঘটে। বিভিন্ন অঙ্গের পেশী তন্তুগুলির সংকোচন এবং শিথিলকরণ নিয়ন্ত্রণের বিভিন্ন আণবিক প্রক্রিয়া থাকতে পারে তবে Ca 2+ আয়ন সর্বদা মূল নিয়ন্ত্রক ভূমিকা পালন করে। এটি প্রতিষ্ঠিত হয়েছে যে মায়োফাইব্রিলগুলির ATP-এর সাথে যোগাযোগ করার ক্ষমতা রয়েছে এবং শুধুমাত্র মাধ্যমের ক্যালসিয়াম আয়নের নির্দিষ্ট ঘনত্বের উপস্থিতিতে এটির উপস্থিতিতে সংকোচন করতে পারে। সর্বাধিক সংকোচনশীল কার্যকলাপ প্রায় 10-6-10-5 M এর Ca 2+ আয়নগুলির ঘনত্বে পরিলক্ষিত হয়। যখন ঘনত্ব 10-7 M বা তার নিচে নেমে যায়, তখন পেশী তন্তুগুলি উপস্থিতিতে টান বাড়ানোর এবং সংক্ষিপ্ত করার ক্ষমতা হারিয়ে ফেলে। ATP এর।
আধুনিক ধারণা অনুসারে, একটি বিশ্রামের পেশীতে (মায়োফাইব্রিলস এবং ইন্টারফাইব্রিলার স্পেসে), সারকোপ্লাজমিক রেটিকুলামের কাঠামো (টিউবুলস এবং ভেসিকেল) দ্বারা আবদ্ধ হওয়ার ফলে Ca 2+ আয়নের ঘনত্ব থ্রেশহোল্ড মানের নীচে বজায় থাকে। একটি বিশেষ Ca 2+ - বাইন্ডিং প্রোটিনের অংশগ্রহণের সাথে টি-সিস্টেম বলা হয়, যাকে বলা হয় ক্যালসকোয়েস্ট্রিন, যা এই কাঠামোর অংশ।
সারকোপ্লাজমিক রেটিকুলামের টিউবুল এবং সিস্টারনের একটি বিস্তৃত নেটওয়ার্ক দ্বারা Ca 2+ আয়নগুলির বাঁধন একটি সহজ শোষণ নয়। এটি একটি সক্রিয় শারীরবৃত্তীয় প্রক্রিয়া, যা সারকোপ্লাজমিক রেটিকুলামের ATP Ca 2+-নির্ভর ATPase এর ভাঙ্গনের সময় নির্গত শক্তির কারণে সঞ্চালিত হয়। এই ক্ষেত্রে, একটি খুব অদ্ভুত চিত্র পরিলক্ষিত হয়: আন্তঃফাইব্রিলার স্থান থেকে Ca 2+ আয়নগুলিকে বহিষ্কারের হার একই আয়ন দ্বারা উদ্দীপিত হয়। সাধারণভাবে, এই ধরনের প্রক্রিয়াটিকে শারীরবিদ্যায় সুপরিচিত সোডিয়াম পাম্পের সাথে সাদৃশ্য দ্বারা "ক্যালসিয়াম পাম্প" বলা হত (চিত্র 11, বি দেখুন)।
এটিপিতে যথেষ্ট পরিমাণে উচ্চ ঘনত্বের উপস্থিতিতে একটি জীবন্ত পেশী শিথিল অবস্থায় থাকার সম্ভাবনা ক্যালসিয়ামের ক্রিয়াকলাপের ফলে মায়োফাইব্রিলগুলির আশেপাশের পরিবেশে Ca 2+ আয়নগুলির ঘনত্ব হ্রাস দ্বারা ব্যাখ্যা করা হয়। পাম্প, সীমার নীচে যেখানে ATPase কার্যকলাপের প্রকাশ এবং ফাইবারের অ্যাক্টোমায়োসিন কাঠামোর সংকোচন এখনও সম্ভব। পেশী ফাইবারের দ্রুত সংকোচন যখন এটি একটি স্নায়ু দ্বারা উদ্দীপিত হয় (বা বৈদ্যুতিক প্রবাহ দ্বারা) ঝিল্লির ব্যাপ্তিযোগ্যতার আকস্মিক পরিবর্তনের ফলে এবং ফলস্বরূপ, একটি নির্দিষ্ট পরিমাণ Ca 2+ নির্গত হয়। সারকোপ্লাজমিক রেটিকুলামের ট্যাঙ্ক এবং টিউবুল থেকে আয়ন এবং টি-সিস্টেম সারকোপ্লাজমে প্রবেশ করে (চিত্র 11, এ, বি দেখুন)।
যেমন উল্লেখ করা হয়েছে, Ca 2+ আয়নগুলির প্রতি অ্যাক্টোমায়োসিন সিস্টেমের "সংবেদনশীলতা" (অর্থাৎ, Ca 2+ আয়নগুলির ঘনত্ব 10-এ কমে যাওয়ার সাথে ATP-এর উপস্থিতিতে অ্যাক্টোমায়োসিনের ATP ক্লিভ করার ক্ষমতা এবং সংকোচনের ক্ষমতা হারানো। –7 M) সংকোচন পদ্ধতিতে উপস্থিতির কারণে (এফ-অ্যাক্টিনের ফিলামেন্টে) ট্রপোমায়োসিনের সাথে যুক্ত ট্রপোনিন প্রোটিন। ট্রপোনিন-ট্রপোমায়োসিন কমপ্লেক্সে, Ca 2+ আয়নগুলি ট্রপোনিনের সাথে সুনির্দিষ্টভাবে আবদ্ধ হয় (ট্রপোনিনের সি-সাবুনিট ক্যালমোডুল-এর বৈশিষ্ট্যে অনুরূপ।

ভাত। এগারো পেশী সংকোচন নিয়ন্ত্রণ
আমরা হব). Ca 2+ আয়নের বাঁধাই ট্রপোনিন অণুতে গঠনমূলক পরিবর্তন ঘটায়, যা দৃশ্যত, পুরো ট্রোপোনিন-ট্রপোমায়োসিন রডের পরিবর্তন ঘটায় এবং অ্যাক্টিন সক্রিয় সাইটগুলিকে অবরুদ্ধ করে যা মায়োসিনের সাথে যোগাযোগ করতে পারে একটি সংকোচনশীল জটিল এবং একটি সক্রিয়। Mg 2+ -ATPase. এটি পেশী সংকোচনের একটি চক্র শুরু করে (চিত্র 11বি দেখুন)।
মায়োসিন ফিলামেন্টের সাথে অ্যাক্টিন ফিলামেন্টের প্রচারে, ই. হাক্সলির মতে, একটি গুরুত্বপূর্ণ ভূমিকা পালন করে ট্রান্সভার্স ব্রিজগুলি অস্থায়ীভাবে ফিলামেন্টগুলির মধ্যে বন্ধ হয়ে যায়, যা মায়োসিন অণুর "মাথা"। সুতরাং, একটি নির্দিষ্ট মুহুর্তে অ্যাক্টিন ফিলামেন্টের সাথে সংযুক্ত সেতুর সংখ্যা যত বেশি, পেশী সংকোচনের শক্তি তত বেশি।
অবশেষে, যদি উত্তেজনা বন্ধ হয়ে যায়, সারকোপ্লাজমে Ca 2+ আয়নের বিষয়বস্তু হ্রাস পায় (ক্যালসিয়াম পাম্প), ফলস্বরূপ, ট্রপোনিন সি সহ Ca 2+ কমপ্লেক্স বিচ্ছিন্ন হয়ে যায়, ট্রপোনিন তার আসল রূপ পুনরুদ্ধার করে, অ্যাক্টিনের মায়োসিন বাইন্ডিং সাইট অবরুদ্ধ হয়। , অর্থাৎ, মায়োসিন ফিলামেন্টের "মাথা" অ্যাক্টিন ফিলামেন্টের সাথে সংযুক্ত হওয়া বন্ধ করে। এটিপির উপস্থিতিতে, পেশী শিথিল হয় এবং এর দৈর্ঘ্য তার আসল দৈর্ঘ্যে পৌঁছায়। যদি এটিপি সরবরাহ বন্ধ হয়ে যায় (অ্যানোক্সিয়া, ইনহেলেশন বিষক্রিয়া, বা মৃত্যু), তাহলে পেশী কঠোর অবস্থায় চলে যায়। মোটা (মায়োসিন) ফিলামেন্টের প্রায় সব ট্রান্সভার্স ব্রিজ পাতলা অ্যাক্টিন ফিলামেন্টের সাথে সংযুক্ত থাকে, যার ফলে পেশীর সম্পূর্ণ অচলতা দেখা দেয়।
অধ্যায় 4. পেশী কার্যকলাপের বায়োএনার্জি
4.1. সিস্টেম এবং মেকানিজমের সাধারণ বৈশিষ্ট্য
পেশী কার্যকলাপের শক্তি সরবরাহ
অধ্যায় 3 এ দেখানো হয়েছে, এটিপি পেশী কার্যকলাপের জন্য শক্তির তাত্ক্ষণিক উৎস। ADP এবং অর্থোফসফেটে ATP অণুর এনজাইমেটিক হাইড্রোলাইসিসের সময় শক্তির মুক্তি ঘটে:
Ca 2+ -ATPase
ATP + H 2 O ADP + H 3 RO 4।
ΔQ = 7.3 kcal, বা 30 kJ
পেশী সংকোচনের প্রক্রিয়ায় রাসায়নিক শক্তি পেশীগুলির যান্ত্রিক কাজে রূপান্তরিত হয় এবং শিথিল হওয়ার সময় এটি সারকোপ্লাজমিক রেটিকুলামে Ca 2+ এর সক্রিয় পরিবহন সরবরাহ করে। Na + -K + -ATPase এর কাজের জন্য কঙ্কালের পেশীগুলিতে প্রচুর পরিমাণে ATP খাওয়া হয়, যা পেশীতে Na + এবং K + আয়নগুলির একটি নির্দিষ্ট ঘনত্ব বজায় রাখে, সারকোলেমাতে একটি ইলেক্ট্রোকেমিক্যাল সম্ভাবনা তৈরি করে।
সুতরাং, এটিপি আকারে পর্যাপ্ত পরিমাণ শক্তি সহ একটি পেশী কোষকে তার সংকোচনযন্ত্রের সাথে সরবরাহ করার জন্য, এই যৌগটির অবিচ্ছিন্ন পুনঃসংশ্লেষণ প্রয়োজন।
পেশীতে ATP-এর বিষয়বস্তু নগণ্য এবং কাঁচা টিস্যু ভরের (0.25–0.40%) প্রায় 5 mmol ∙ kg -1। এটি তুলনামূলকভাবে ধ্রুবক স্তরে রক্ষণাবেক্ষণ করা হয়, যেহেতু পেশীতে ATP-এর ঘনত্ব বৃদ্ধির ফলে মায়োসিন ATPase-কে বাধা দেয়, যা মায়োফাইব্রিল এবং পেশী সংকোচনে অ্যাক্টিন এবং মায়োসিন ফিলামেন্টের মধ্যে আনুগত্য গঠনে বাধা দেয় এবং এটি 2 mmol ∙ এর নিচে হ্রাস পায়। কাঁচা টিস্যু ভরের কেজি -1 জালিকায় Ca 2+ পাম্পের কাজ এবং পেশী শিথিলকরণের প্রক্রিয়া লঙ্ঘনের দিকে পরিচালিত করে। পেশী ফাইবারগুলিতে ATP মজুদ শুধুমাত্র খুব অল্প সময়ের জন্য তীব্র কাজের কার্যকারিতা নিশ্চিত করতে পারে - 0.5-1.5 সেকেন্ড, বা সর্বাধিক শক্তির 3-4 একক সংকোচন। এটিপির ক্ষয়ের পণ্যগুলি থেকে দ্রুত পুনরুদ্ধার (পুনঃসংশ্লেষণ) এবং ক্ষয়ের সময় যে পরিমাণ শক্তি নির্গত হয়েছিল তার কারণে আরও পেশীর কাজ করা হয়:
ADP + H 3 RO 4 + ΔQ → ATP।
ফসফেট যোগ করার বিক্রিয়াকে বলা হয় ফসফোরিলেশন, এবং একে এক পদার্থ থেকে অন্য পদার্থে স্থানান্তরের বিক্রিয়াকে বলা হয় রেফসফোরিলেশন।


ভাত। 12. পেশী টিস্যুতে শক্তি বিপাক
কঙ্কালের পেশী এবং অন্যান্য টিস্যুতে এটিপি পুনঃসংশ্লেষণের জন্য শক্তির উত্সগুলি হল শক্তি সমৃদ্ধ ফসফেটযুক্ত পদার্থ যা টিস্যুতে উপস্থিত থাকে (ক্রিয়েটাইন ফসফেট, এডিপি) বা গ্লাইকোজেন, ফ্যাটি অ্যাসিড এবং অন্যান্য শক্তির স্তরগুলির (উদাহরণস্বরূপ, মেটাবোলাইটস) এর বিপাক করার সময় গঠিত হয় diphosphoglyceric এবং phosphopyruvic অ্যাসিড), এবং এছাড়াও বিভিন্ন পদার্থের বায়বীয় জারণের ফলে মাইটোকন্ড্রিয়াল ঝিল্লি জুড়ে প্রোটন (H +) গ্রেডিয়েন্টের শক্তি।
কোন জৈব রাসায়নিক প্রক্রিয়া এটিপি অণু গঠনের জন্য শক্তি সরবরাহ করে তার উপর নির্ভর করে, টিস্যুতে এটিপি পুনঃসংশ্লেষণের জন্য চারটি প্রক্রিয়া বা পথ রয়েছে (চিত্র 12 দেখুন)। প্রতিটি প্রক্রিয়ার নিজস্ব বিপাকীয় এবং বায়োএনার্জেটিক বৈশিষ্ট্য রয়েছে। পেশীবহুল কাজের শক্তি সরবরাহে, অনুশীলনের তীব্রতা এবং সময়কালের উপর নির্ভর করে বিভিন্ন প্রক্রিয়া ব্যবহার করা হয়।
এটিপি পুনঃসংশ্লেষণ অক্সিজেন (অ্যানেরোবিক প্রক্রিয়া) ব্যতীত বা ইনহেলড অক্সিজেন (বায়ুবিক প্রক্রিয়া) এর অংশগ্রহণ ছাড়াই ঘটে এমন প্রতিক্রিয়াগুলিতে সঞ্চালিত হতে পারে।
স্বাভাবিক অবস্থায়, টিস্যুতে এটিপি সংশ্লেষণ প্রধানত বায়বীয়ভাবে ঘটে এবং তীব্র পেশী ক্রিয়াকলাপের সময়, যখন পেশীতে অক্সিজেন সরবরাহ করা কঠিন হয়, টিস্যুতে এটিপি সংশ্লেষণের অ্যানেরোবিক প্রক্রিয়াও বৃদ্ধি পায়। মানুষের কঙ্কালের পেশীতে, তিন ধরনের অ্যানেরোবিক এবং এটিপি পুনর্সংশ্লেষণের একটি বায়বীয় পথ চিহ্নিত করা হয়েছে (চিত্র 13 দেখুন)।
অ্যানেরোবিক প্রক্রিয়া অন্তর্ভুক্ত:
1) ক্রিয়েটাইন ফসফোকিনেস (ফসফোজেনিক বা অ্যাল্যাক্টেট) প্রক্রিয়া যা ক্রিয়েটাইন ফসফেট এবং এডিপির মধ্যে রিফসফোরিলেশনের কারণে এটিপি রিসিন্থেসিস প্রদান করে;
2) গ্লাইকোলাইটিক (ল্যাকটেট) প্রক্রিয়া, যা পেশী গ্লাইকোজেন বা রক্তের গ্লুকোজের এনজাইম্যাটিক অ্যানেরোবিক ভাঙ্গনের প্রক্রিয়ায় এটিপি পুনঃসংশ্লেষণ প্রদান করে, ল্যাকটিক অ্যাসিড গঠনের সাথে শেষ হয় এবং তাই ল্যাকটেট বলা হয়;
3) মায়োকিনেস মেকানিজম, যা এনজাইম মায়োকিনেজ (অ্যাডিনাইলেট কাইনেস) এর অংশগ্রহণের সাথে দুটি ADP অণুর মধ্যে রিফোসফোরিলেশন প্রতিক্রিয়ার কারণে এটিপিকে পুনরায় সংশ্লেষণ করে।

ভাত। 13. পেশীতে এটিপি সংশ্লেষণের প্রক্রিয়া
(ফ্রেমওয়ার্ক শক্তি সাবস্ট্রেট দেখায়
এবং মেকানিজমের নাম হাইলাইট করেছেন)
এটিপি পুনঃসংশ্লেষণের বায়বীয় প্রক্রিয়ায় প্রধানত মাইটোকন্ড্রিয়ায় অক্সিডেটিভ ফসফোরিলেশন প্রতিক্রিয়া অন্তর্ভুক্ত থাকে। অ্যারোবিক অক্সিডেশনের শক্তির স্তরগুলি হল গ্লুকোজ, ফ্যাটি অ্যাসিড, আংশিকভাবে অ্যামিনো অ্যাসিড, পাশাপাশি গ্লাইকোলাইসিসের মধ্যবর্তী বিপাক - ল্যাকটিক অ্যাসিড, ফ্যাটি অ্যাসিড অক্সিডেশন - কেটোন বডি।
প্রতিটি প্রক্রিয়ার বিভিন্ন শক্তি ক্ষমতা রয়েছে, যা শক্তি উৎপাদন প্রক্রিয়া মূল্যায়নের জন্য নিম্নলিখিত মানদণ্ড দ্বারা চিহ্নিত করা হয়: সর্বাধিক শক্তি, স্থাপনার হার, বিপাকীয় ক্ষমতা এবং দক্ষতা। সর্বোচ্চ শক্তি হল একটি প্রদত্ত বিপাকীয় প্রক্রিয়ায় এটিপি উৎপাদনের সর্বোচ্চ হার। এটি এই প্রক্রিয়া দ্বারা সম্পাদিত কাজের সর্বাধিক তীব্রতা সীমাবদ্ধ করে। কাজের শুরু থেকে প্রদত্ত ATP রেসিন্থেসিস পাথওয়ের সর্বোচ্চ শক্তিতে পৌঁছানোর সময় দ্বারা স্থাপনার হার অনুমান করা হয়। বিপাকীয় ক্ষমতা শক্তি সাবস্ট্রেটের রিজার্ভের আকারের কারণে একটি প্রদত্ত রেসিন্থেসিস প্রক্রিয়ায় প্রাপ্ত ATP-এর মোট পরিমাণ প্রতিফলিত করে; ক্ষমতা কাজ করা যেতে পারে যে পরিমাণ সীমিত. বিপাকীয় দক্ষতা হল শক্তির সেই অংশ যা ATP-এর ম্যাক্রোঅার্জিক বন্ডে জমা হয়; এটি সম্পাদিত কাজের দক্ষতা নির্ধারণ করে এবং কার্যক্ষমতা সহগ (COP) এর সামগ্রিক মূল্য দ্বারা অনুমান করা হয়, যা এই বিপাকীয় প্রক্রিয়ায় মুক্তির মোট পরিমাণের সাথে ব্যয় করা সমস্ত দরকারী শক্তির অনুপাত।
বিপাকীয় প্রক্রিয়ার শক্তিকে যান্ত্রিক কাজে (Em) রূপান্তর করার সামগ্রিক দক্ষতা দুটি সূচকের উপর নির্ভর করে: ক) বিপাকীয় রূপান্তরের সময় মুক্তি পাওয়া শক্তিকে পুনরায় সংশ্লেষিত ম্যাক্রোএনার্জেটিক ফসফরাস যৌগগুলির (এটিপি) শক্তিতে রূপান্তর করার দক্ষতা, অর্থাৎ, ফসফোরিলেশনের দক্ষতা (E f); খ) এটিপিকে যান্ত্রিক কাজে রূপান্তর করার দক্ষতা, অর্থাৎ কেমোমেকানিক্যাল কাপলিং এর দক্ষতা (ই ই):
E m = (E f/E e) × 100।
অ্যারোবিক এবং অ্যানেরোবিক বিপাকের শতাংশ হিসাবে কেমোমেকানিক্যাল কনজুগেশনের কার্যকারিতা প্রায় একই এবং এর পরিমাণ 50%, যখন ফসফোরিলেশন কার্যকারিতা অ্যাল্যাকটিক অ্যানেরোবিক প্রক্রিয়াতে সর্বোচ্চ - প্রায় 80% এবং সর্বনিম্ন - অ্যানেরোবিক গ্লাইকোর গড়। 44%, বায়বীয় প্রক্রিয়ায় এটি প্রায় 60%।
মূল্যায়নের মানদণ্ড অনুসারে পেশী কার্যকলাপের শক্তি সরবরাহের ক্রিয়েটাইন ফসফোকিনেস, গ্লাইকোলাইটিক এবং বায়বীয় প্রক্রিয়ার তুলনামূলক বৈশিষ্ট্যগুলি সারণি 3 এ উপস্থাপন করা হয়েছে।
সারণি 3 দেখায় যে ক্রিয়েটাইন ফসফোকিনেস এবং গ্লাইকোলাইটিক প্রক্রিয়াগুলির ATP গঠনের একটি বৃহৎ সর্বাধিক শক্তি এবং দক্ষতা রয়েছে, তবে একটি সংক্ষিপ্ত সর্বাধিক শক্তি ধারণ সময় এবং শক্তির স্তরগুলির ক্ষুদ্র মজুদের কারণে একটি ছোট ক্ষমতা। ক্রিয়েটাইন ফসফোকিনেসের তুলনায় অ্যারোবিক মেকানিজমের সর্বোচ্চ শক্তি প্রায় তিনগুণ কম, তবে এটি দীর্ঘ সময়ের জন্য বজায় রাখে, সেইসাথে কার্বোহাইড্রেট, চর্বি এবং আংশিকভাবে প্রোটিনের আকারে শক্তির স্তরগুলির বিশাল মজুদের কারণে প্রায় অক্ষয় ক্ষমতা। সুতরাং, চর্বিগুলির মজুদের কারণে, শরীর ক্রমাগত 7-10 দিন কাজ করতে পারে, যখন শক্তি উত্পাদনের অ্যানেরোবিক প্রক্রিয়াগুলির শক্তি সাবস্ট্রেটগুলির মজুদ কম তাৎপর্যপূর্ণ।
প্রশিক্ষণ নথি
... ইউডিসি (470)(082) বিবিসি ... প্রকাশনাদাতব্য এম.পি. বেলিয়ায়েভ। স্বতন্ত্র দল প্রকাশনা ... মুদ্রিত ... উপদেশযেখানে এটি গৃহীত হয়েছিল সমাধানপ্রবাসে তাদের কার্যক্রম চালিয়ে যান। মায়ায় লিপ্ত নয় চালু... গুণমান শিক্ষাগত সুবিধা. তার...