Кои органи съдържат мускулна тъкан. Функции на гладката мускулна тъкан
Подробности
ганглиипредставлявам клъстери от мултиполярни (един аксон и няколко дендрита) неврони(от няколко клетки до десетки хиляди). Екстраорганните (симпатикови) ганглии имат добре изразена съединителнотъканна капсула, като продължение на периневриума. Парасимпатиковите ганглии обикновено се намират в интрамуралните нервни плексуси. Ганглиите на интрамуралните плексуси, подобно на други автономни възли, съдържат автономни неврони на локални рефлексни дъги. Мултиполярните неврони с диаметър 20-35 μm са разположени дифузно, всеки неврон е заобиколен от ганглийни глиоцити.
Освен това описано невроендокринни, хеморецепторни, биполярни и при някои гръбначни животни, униполярни неврони. В симпатиковите ганглии има малки интензивно флуоресцентни клетки (MYF клетки) с къси процеси и голям брой гранулирани везикули в цитоплазмата. Те секретират катехоламини и имат инхибиторен ефект върху предаването на импулси от преганглионарните нервни влакна към еферентния симпатиков неврон. Тези клетки се наричат интерневрони.
Сред основните мултиполярни невронивегетативен ганглииразграничават: моторни (клетки на Догел от 1-ви тип), чувствителни (клетки на Догел от II-ти тип) и асоциативни (клетки на Догел от III-ти тип). Моторните неврони имат къси дендрити с ламеларни разширения („рецептивни подложки“). Аксонът на тези клетки е много дълъг, надхвърля ганглия като част от постганглионарните тънки немиелинизирани нервни влакна и завършва на гладки миоцити на вътрешните органи. Клетките от 1-ви тип се наричат неврони с дълъг аксон. Невроните от II тип са равноотдалечени нервни клетки. От тялото им излизат 2-4 процеса, сред които е трудно да се различи аксон. Без да се разклоняват, процесите отиват далеч от тялото на неврона. Техните дендрити имат чувствителни нервни окончания, а аксонът завършва върху телата на двигателните неврони в съседните ганглии. Клетките тип II са чувствителни неврони на локалните автономни рефлексни дъги. Тип III Dogel клетки са подобни по форма на тялото на тип II автономни неврони, но техните дендрити не се простират отвъд ганглия и невритът отива към други ганглии. Много изследователи смятат тези клетки за разновидности на чувствителни неврони.
По този начин в периферните автономни ганглии има локални рефлексни дъги, състоящи се от сензорни, двигателни и евентуално асоциативни автономни неврони.
Интрамуралните автономни ганглии в стената на храносмилателния тракт се отличават с факта, че в допълнение към моторните холинергични неврони съдържат инхибиторни неврони. Те са представени от адренергични и пуринергични нервни клетки. В последния медиаторът е пуринов нуклеотид. В интрамуралните автономни ганглии има и пептидергични неврони, които секретират вазоинтестинален пептид, соматостатин и редица други пептиди, с помощта на които се извършва невроендокринна регулация и модулиране на активността на тъканите и органите на храносмилателната система.
Ацетилхолин- никотинови (блок на кураре, хексаметоний), мускаринови (блок на атропин) рецептори. Рецепторно активиране → генериране на EPSP. Бърз EPSP (N-холинергичен)→отваряне на йонни канали. Бавен EPSP (М-холинергичен) → потискане на М-ток поради повишаване на К-проводимостта.
Невропептиди- действат като невромодулатори.
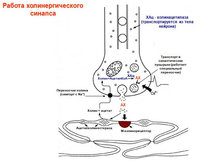
Енкефалини, субстанция Р, люлиберин, невротензин, соматостатин - симптом. ганглии (+Ach)
Катехоламини(NA, допамин) са дребноклетъчни невротрансмитери с интензивна флуоресценция.
Невропептид Y, соматостатин - симптом. постганглионарна.
Симпатични постганглионарни клетки: NA, ATP, невропептид U.
α1→инозотолов трифосфат, диацилглицерол. α2→G-протеин активиране, ↓cAMP.
β→G-протеин→AC→cAMP

Изключения: медиатор Ach, мускаринови рецептори.
парасимп. постганглионарни: Ach, VIP, NO, соматостатин, ATP, опиоидни пептиди.
М1 (висок афинитет към пирензепин) - повишена киселинна секреция от клетките на жлезите на стомаха, М2 (нисък) - забавяне на сърдечната дейност. ритъм, секреция на слъзните и слюнчените жлези.
Разни действия:
-Конкретен сек. медиатори: M2 може да действа на IP3 или може да индуцира AC, намалявайки cAMP.
- Действие върху K и Ca каналите
- NO → гуанилатциклаза → cGMP → cGMP-зависима протеин киназа → върху ендотела се образува гладкомускулна релаксация.
Сензорните ганглии съдържат псевдоунипорални или биполярни аферентни неврони и са разположени по протежение на задните коренчета на гръбначния мозък (гръбначни или спинални ганглии) и черепните нерви (V, VII, VIII, IX, X).
гръбначен възел (ganglion spinale)покрити със съединителнотъканна капсула. Вътре в възела има групи от псевдо-униполярни сензорни неврони, между които преминават снопове от миелинови влакна. Цитоплазмата на клетките съдържа множество митохондрии, rREPS цистерни, комплекс Голджи и лизозоми. Телата на невроните са заобиколени от клетки - сателити(клетки на мантията) и съединителнотъканна капсула. Ганглийните неврони са три вида: малки, междинни и големи. Те се различават по видовете импулси, които провеждат (тактилна чувствителност, пропрецепция, болка, предават информация за мускулната дължина и мускулния тонус на централната нервна система и др.). Те съдържат невротрансмитери: субстанция Р, соматостатин и холецистокинин, глутамин, VIP, гастрин. Техните периферни процеси завършват в периферията с рецептори. Централните процеси (аксони) навлизат в гръбначния мозък, образувайки задните корени на гръбначния мозък и завършват в синапси на интерневроните и моторните неврони на предните рога на гръбначния мозък.
Автономни (вегетативни) нервни възли (ганглии) разположени по дължината на гръбначния стълб под формата на верига (паравентебрални ганглии) и пред него (превентебрални ганглии), както и в стената на органите - сърцето, храносмилателния тракт, пикочния мехур и др. (интрамурални ганглии) или близо до повърхността на органите (екстрамурални ганглии) .
към автономните ганглииподходящи преганглионарни влакна (миелин, съдържащ процеси на нервни клетки, телата на които лежат в централната нервна система. Влакната се разклоняват силно и образуват синаптични окончания върху клетките на автономните ганглии.
Автономни ганглии на функционална основа и локализация се делят на симпатикова и парасимпатикова.
Симпатични ганглии (паравертебрални и превертебрални)получават преганглионарни влакна от клетки, разположени в автономните ядра на гръдния и лумбалния сегмент на гръбначния мозък. Невротрансмитерът на преганглионарните влакна е ацетилхолин, апостганглионарна - норепинефрин (с изключение на потните жлези и някои кръвоносни съдове, които имат хилинергична симпатикова инервация) Енкефалини, VIP, субстанция Р, соматостатин, холецистокенин също се откриват във възлите.
Парасимпатикови ганглии(интрамурални, екстрамурални или възли на главата) получават преганглионарни влакна от клетки, чиито тела са разположени в автономните ядра на продълговатия мозък, средния мозък и в сакралния гръбначен мозък. Тези влакна напускат ЦНС като част от III, VII, IX и X двойки черепни нерви и предните корени на сакралните сегменти на гръбначния мозък. Невротрансмитерът на пре- и постганглионарните влакна е ацетилхолин, както и серотонин АТФ и др.
Повечето органи получават както симпатикова, така и парасимпатикова инервация.
Структурата на симпатиковата и парасимпатиковатаганглиите като цяло са подобни. Вегетативните ганглии от повърхността са покрити със съединителнотъканна капсула, която, прониквайки в възела, образува строма. Възлите се състоят от многополюсни нервни клетки, различни по форма и размери и техните процеси. Телата на неврони с неправилна форма, с ексцентрично разположени ядра, са заобиколени от черупки на глиални клетки - сателити (мантийни глиоцити). Израстъците на клетките също са покрити с глиални клетки. Глиалната обвивка е покрита от базална мембрана, върху която има обвивка от съединителна тъкан.
Симпатиковите ганглии, заедно с големите клетки, съдържат малки групи от малки клетки с интензивно флуоресцентни гранули, MIF клетки и малки клетки, съдържащи гранули (MSH клетки). Гранулите съдържат допамин, серотонин или норепинефрин. Терминалите на преганглионарните влакна завършват на MIF клетки, при стимулиране на които медиаторите се освобождават в периваскуларните пространства и в областта на синапсите на дендритите на големи клетки. MYTH клетките имат инхибиращ ефект върху ефекторните клетки.
интрамурални възли- това са нервни ганглии, разположени вътре в инервираните органи. Интрамуралните ганглии и пътищата, свързани с тях, имат висока автономност, сложност на организацията и особеностите на медиаторите и в тази връзка много автори ги разграничават като независим метасимпатиков отдел на автономната нервна система.
Структурата на интрамуралните ганглии може да се види най-ясно на примера на автономната инервация на храносмилателния тракт. Храносмилателната тръба съдържа два големи нервни плексуса: субмукозен - Майснер, междумускулно - Ауербах.Общият брой на невроните в интрамуралните ганглии е по-висок от този в гръбначния мозък и по отношение на сложността на тяхното взаимодействие те се сравняват с микрокомпютър.
В интрамуралните възли са описани неврони от 3 вида. Първите данни за хетерогенността на невроните в храносмилателния тракт са получени от Догел. Въз основа на формата на клетките и естеството на разклоняването на техните израстъци Догел идентифицира три вида неврони.
1. еферентни неврони с дълъг аксон (клетки на Dogel тип I) преобладават числено. Това са големи или средни клетки, сплескани перикарии с къси дендрити и дълъг аксон, който излиза извън възела и завършва върху клетките на работния орган с двигателен или секреторен край.
Това е система от тъкани и органи, изградени от нервна тъкан. Той подчертава:
Централна област: мозък и гръбначен мозък
Периферни: автономни и сензорни ганглии, периферни нерви, нервни окончания.
Има и разделение на:
Соматично (анимално, цереброспинално) отделение;
Вегетативен (автономен) отдел: симпатикови и парасимпатикови части.
Нервната система се формира от следните ембрионални източници: неврална тръба, нервен гребен (ганглиозна плоча) и ембрионални плакоди. Тъканните елементи на мембраните са мезенхимни производни. На етапа на затваряне на невропорите предният край на тръбата се разширява значително, страничните стени се удебеляват, образувайки началото на три церебрални везикули. Пикочният мехур, разположен краниално, образува предния мозък, средният пикочен мехур образува средния мозък, а задният (ромбоиден) мозък се развива от третия пикочен мехур, който преминава в залата на гръбначния мозък. Скоро след това невралната тръба се огъва почти под прав ъгъл и чрез стесняващите се бразди първият пикочен мехур се разделя на краен и междинен отдел, а третият мозъчен мехур - на продълговатия мозък и задните отдели на мозъка. Производните на средните и задните мозъчни везикули образуват мозъчния ствол и са древни образувания; те запазват сегментния принцип на структурата, който изчезва в производните на диенцефалона и теленцефалона. В последния са съсредоточени интегративни функции. Така се образуват пет части на мозъка: краен и диенцефалон, среден, продълговат и заден мозък (при хората това се случва приблизително в края на 4-та седмица от ембрионалното развитие). Теленцефалонът образува двете полукълба на главния мозък.
В ембрионалната хисто- и органогенеза на нервната система развитието на различни части на мозъка протича с различна скорост (хетерохронно). По-рано се формират каудалните части на централната нервна система (гръбначен мозък, мозъчен ствол); времето на окончателното формиране на мозъчните структури варира значително. В редица части на мозъка това се случва след раждането (малък мозък, хипокампус, обонятелна луковица); във всяка част на мозъка има пространствено-времеви градиенти при формирането на невронни популации, които образуват уникална структура на нервния център.
Гръбначният мозък е част от централната нервна система, в чиято структура са най-ясно запазени характеристиките на ембрионалните етапи на развитие на мозъка на гръбначните животни: тръбният характер на структурата и сегментацията. В страничните участъци на невралната тръба масата на клетките бързо нараства, докато нейните дорзални и вентрални части не се увеличават по обем и запазват своя епендимален характер. Удебелените странични стени на невралната тръба са разделени от надлъжен жлеб на дорзална - аларна и вентрална - основна плоча. На този етап на развитие могат да се разграничат три зони в страничните стени на невралната тръба: епендима, облицоваща централния канал, междинна (слой на мантията) и маргинална (маргинален воал). Сивото вещество на гръбначния мозък впоследствие се развива от мантийния слой, а бялото му вещество се развива от маргиналния воал. Невробластите на предните колони се диференцират в моторни неврони (моторни неврони) на ядрата на предните рога. Техните аксони излизат от гръбначния мозък и образуват предните коренчета на гръбначномозъчните нерви. В задните колони и междинната зона се развиват различни ядра от интеркаларни (асоциативни) клетки. Техните аксони, влизащи в бялото вещество на гръбначния мозък, са част от различни проводящи снопове. Задните рога включват централните процеси на сетивните неврони на гръбначните възли.
Едновременно с развитието на гръбначния мозък се полагат гръбначните и периферните възли на автономната нервна система. Изходен материал за тях са елементите на стволовите клетки на нервния гребен, които чрез дивергентна диференциация се развиват в невробластно и глиобластично направление. Част от клетките на нервния гребен мигрират към периферията към местата на локализация на възлите на автономната нервна система, параганглиите, невроендокринните клетки от серията APUD и хромафиновата тъкан.
Периферна нервна система.
Периферната нервна система съчетава периферни нервни възли, стволове и окончания.
Нервните ганглии (възли) - структури, образувани от клъстери от неврони извън централната нервна система - се разделят на чувствителни и автономни (вегетативни). Сензорните ганглии съдържат псевдо-униполярни или биполярни (в спиралните и вестибуларните ганглии) аферентни неврони и са разположени главно по протежение на задните корени на гръбначния мозък (сензорни възли на гръбначните нерви) и някои черепни нерви. Сетивните ганглии на гръбначномозъчните нерви са вретеновидни и покрити с капсула от плътна фиброзна съединителна тъкан. По периферията на ганглия има плътни клъстери от тела на псевдоуниполярни неврони, а централната част е заета от техните процеси и тънки слоеве ендоневриум, разположени между тях, носещи съдове. Автономните нервни ганглии се образуват от клъстери от мултиполярни неврони, върху които множество синапси образуват преганглионарни влакна - процеси на неврони, чиито тела лежат в ЦНС.
нерв. Изграждане и регенерация. Спинални ганглии. Морфофункционални характеристики.
Нервите (нервните стволове) свързват нервните центрове на главния и гръбначния мозък с рецептори и работни органи. Те се образуват от снопове миелинизирани и немиелинизирани влакна, които са обединени от компоненти на съединителната тъкан (черупки): ендоневриум, периневриум и епиневриум. Повечето от нервите са смесени, т.е. включват аферентни и еферентни влакна.
Ендоневриум - тънки слоеве от рехава влакнеста съединителна тъкан с малки кръвоносни съдове, обграждащи отделни нервни влакна и свързващи ги в един сноп. Периневриумът е обвивка, която покрива всеки сноп от нервни влакна отвън и разширява преградите дълбоко в снопа. Има ламеларна структура и изображения на концентрични слоеве от сплескани фибробластоподобни клетки, свързани с плътни и нарязани стави. Между слоевете клетки в пространствата, пълни с течност, има компоненти на базалната мембрана и надлъжно ориентирани колагенови влакна. Епиневриумът е външната обвивка на нерва, която свързва снопове от нервни влакна заедно. Състои се от плътна фиброзна съединителна тъкан, съдържаща мастни клетки, кръвоносни и лимфни съдове.
Гръбначен мозък. Морфофункционални характеристики. развитие. Структура на сивото и бялото вещество. нервен състав.
Гръбначният мозък се състои от две симетрични половини, разделени една от друга отпред от дълбока средна фисура и отзад от съединителнотъканна преграда. Вътрешната част на органа е по-тъмна - това е сивото му вещество. По периферията на гръбначния мозък има по-светло бяло вещество. Сивото вещество на гръбначния мозък се състои от тела на неврони, немиелинизирани и тънки миелинизирани влакна и невроглия. Основният компонент на сивото вещество, което го отличава от бялото, са мултиполярните неврони. Издатините на сивото вещество се наричат рога. Има предни или вентрални, задни или гръбни и странични или странични рога. По време на развитието на гръбначния мозък от невралната тръба се образуват неврони, групирани в 10 слоя или в пластини. Характеристика на човек
следната архитектоника на посочените плочи: плочи I-V съответстват на задните рога, плочи VI-VII - на междинната зона, плочи VIII-IX - на предните рога, плоча X - на зоната на близкия централен канал. Сивото вещество на мозъка се състои от три вида мултиполярни неврони. Първият тип неврони е филогенетично по-стар и се характеризира с няколко дълги, прави и слабо разклонени дендрити (изодендритен тип). Вторият тип неврони има голям брой силно разклонени дендрити, които се преплитат, образувайки "заплетения" (идиодендритичен тип). Третият тип неврони, по отношение на степента на развитие на дендритите, заема междинна позиция между първия и втория тип. Бялото вещество на гръбначния мозък е колекция от надлъжно ориентирани предимно миелинизирани влакна. Сноповете нервни влакна, които комуникират между различни части на нервната система, се наричат пътища на гръбначния мозък.
мозък. Източници на развитие. Обща морфофункционална характеристика на мозъчните полукълба. Невронна организация на мозъчните полукълба. Цито- и миелоархитектоника на кората на главния мозък. Свързани с възрастта промени в кората.
В мозъка се разграничават сивото и бялото вещество, но разпределението на тези два компонента тук е много по-сложно, отколкото в гръбначния мозък. По-голямата част от сивото вещество на мозъка е разположено на повърхността на големия мозък и в малкия мозък, образувайки тяхната кора. По-малка част образува множество ядра на мозъчния ствол.
Структура. Кората на главния мозък е представена от слой сиво вещество. Най-силно е развит в предния централен гирус. Изобилието от бразди и извивки значително увеличава площта на сивото вещество на мозъка .. Различните му части, които се различават една от друга по някои характеристики на местоположението и структурата на клетките (цитоархитектоника), местоположението на влакната (миелоархитектоника) и функционално значение, се наричат полета. Те са места за висш анализ и синтез на нервни импулси. рязко дефиниран
между тях няма граници. Кортексът се характеризира с подреждането на клетките и влакната на слоеве. Развитието на човешкия церебрален кортекс (неокортекс) в ембриогенезата възниква от вентрикуларната зародишна зона на теленцефалона, където се намират слабо специализирани пролифериращи клетки. Неокортикалните невроцити се диференцират от тези клетки. В този случай клетките губят способността си да се делят и да мигрират към възникващата кортикална плоча. Първо, невроцитите на бъдещите слоеве I и VI навлизат в кортикалната плоча, т.е. най-повърхностните и дълбоки слоеве на кората. След това в него се вграждат неврони от слоеве V, IV, III и II в посока отвътре и навън. Този процес се осъществява поради образуването на клетки в малки области на вентрикуларната зона в различни периоди на ембриогенезата (хетерохронно). Във всяка от тези области се образуват групи от неврони, последователно подредени по едно или повече влакна.
радиална глия под формата на колона.
Цитоархитектоника на кората на главния мозък.Мултиполярните неврони на кората са много разнообразни по форма. Сред тях са пирамидални, звездовидни, вретеновидни, паякообразни и хоризонтални неврони. Невроните на кората са разположени в неясно разграничени слоеве. Всеки слой се характеризира с преобладаването на който и да е тип клетки. В двигателната зона на кората се разграничават 6 основни слоя: I - молекулен, II - външен гранулиран, III - нурамидни неврони, IV - вътрешен гранулиран, V - ганглийен, VI - слой от полиморфни клетки. Молекулярният слой на кората съдържа малък брой малки вретеновидни асоциативни клетки. Техните неврити вървят успоредно на повърхността на мозъка като част от тангенциалния плексус на нервните влакна на молекулярния слой. Външният гранулиран слой се формира от малки неврони, които имат заоблена, ъглова и пирамидална форма, и звездовидни невроцити. Дендритите на тези клетки се издигат в молекулярния слой. Невритите или отиват в бялото вещество, или, образувайки дъги, също навлизат в тангенциалния плексус на влакната на молекулярния слой. Най-широкият слой на кората на главния мозък е пирамидалният. От върха на пирамидалната клетка се отклонява основният дендрит, който се намира в молекулярния слой. Невритът на пирамидната клетка винаги се отклонява от основата си. Вътрешният гранулиран слой се формира от малки звездовидни неврони. Състои се от голям брой хоризонтални влакна. Ганглийният слой на кората се формира от големи пирамиди, а областта на прецентралната извивка съдържа гигантски пирамиди.
Слоят от полиморфни клетки се образува от неврони с различна форма.
Миелоархитектоника на кората. Сред нервните влакна на кората на главния мозък могат да се разграничат асоциативни влакна, които свързват отделни части на кората на едно полукълбо, комиссурални влакна, които свързват кората на различни полукълба, и проекционни влакна, както аферентни, така и еферентни, които свързват кората с ядрата на долните части на центр
нервна система.
Възрастови промени. През 1-вата година от живота се наблюдава типизиране на формата на пирамидални и звездовидни неврони, тяхното увеличаване, развитие на дендритни и аксонови арборизации и интраансамбълни връзки по вертикалата. До 3-годишна възраст в ансамблите се разкриват „вложени“ групи от неврони, по-ясно оформени вертикални дендритни снопове и снопове от лъчисти влакна. До 5-6-годишна възраст полиморфизмът на невроните се увеличава; системата от вътрешно-ансамбълови връзки по хоризонталата става по-сложна поради нарастването на дължината и разклоняването на страничните и базалните дендрити на пирамидалните неврони и развитието на страничните терминали на техните апикални дендрити. До 9-10-годишна възраст клетъчните групи се увеличават, структурата на късите аксонни неврони става много по-сложна и мрежата от аксонни колатерали на всички форми на интерневрони се разширява. До 12-14-годишна възраст специализираните форми на пирамидални неврони са ясно маркирани в ансамбли; всички видове интернейрони достигат високо ниво на диференциация. До 18-годишна възраст ансамбловата организация на кората по отношение на основните параметри на нейната архитектоника достига нивото на тази при възрастните.
Малък мозък. Структура и морфофункционални характеристики. Невронен състав на кората на малкия мозък, глиоцити. Интерневронни връзки.
Малък мозък. Това е централният орган на баланса и координацията на движенията. Той е свързан с мозъчния ствол чрез аферентни и еферентни проводящи снопове, които заедно образуват три двойки малкомозъчни стъбла. На повърхността на малкия мозък има много извивки и бразди, които значително увеличават площта му. На разреза се създават бразди и извивки
характерна за малкия мозък картина на "дървото на живота". По-голямата част от сивото вещество в малкия мозък е разположено на повърхността и образува неговата кора. По-малка част от сивото вещество се намира дълбоко в бялото вещество под формата на централни ядра. В центъра на всяка извивка има тънък слой
бяло вещество, покрито със слой сиво вещество - кората. В кората на малкия мозък се разграничават три слоя: външният е молекулярен слой, средният е ганглийният слой или слой от крушовидни неврони, а вътрешният е гранулиран. Ганглийният слой съдържа крушовидни неврони. Те имат неврити, които, напускайки кората на малкия мозък, образуват началната връзка на неговия еферент
спирачни пътища. От крушовидното тяло в молекулярния слой се простират 2-3 дендрита, които проникват в цялата дебелина на молекулярния слой. От основата на телата на тези клетки невритите се отклоняват, преминавайки през гранулирания слой на кората на малкия мозък в бялото вещество и завършвайки върху клетките на малкия мозък. Молекулярният слой съдържа два основни типа неврони: кошничкови и звездовидни. Кошничните неврони са разположени в долната трета на молекулярния слой. Техните тънки дълги дендрити се разклоняват главно в равнина, разположена напречно на гируса. Дългите неврити на клетките винаги преминават през гируса и успоредно на повърхността над крушовидните неврони. Звездовидните неврони лежат над кошничковите клетки и биват два вида. Малките звездовидни неврони са снабдени с тънки къси дендрити и слабо разклонени неврити, които образуват синапси. Големите звездовидни неврони имат дълги и силно разклонени дендрити и неврити. зърнест слой. Първият тип клетки в този слой може да се счита за гранулирани неврони или гранулирани клетки. Клетката има 3-4 къси дендрита,
завършващи в същия слой с крайни разклонения под формата на птича лапа. Невритите на зърнестите клетки преминават в молекулярния слой и в него се разделят на два клона, ориентирани успоредно на повърхността на кората по протежение на гирусите на малкия мозък. Вторият тип клетки в гранулирания слой на малкия мозък са инхибиторни големи звездовидни неврони. Има два вида такива клетки: с къси и дълги неврити. Невроните с къси неврити лежат близо до ганглиозния слой. Техните разклонени дендрити се разпространяват в молекулярния слой и образуват синапси с успоредни влакна - аксони на гранулирани клетки. Невритите се изпращат в гранулирания слой към гломерулите на малкия мозък и завършват в синапси в крайните разклонения на дендритите на гранулираните клетки.
Няколко звездовидни неврони с дълги неврити имат дендрити и неврити, изобилно разклонени в гранулирания слой, простиращи се в бялото вещество. Третият вид клетки са вретеновидни хоризонтални клетки. Имат малко удължено тяло, от което в двете посоки се простират дълги хоризонтални дендрити, завършващи в ганглийния и зърнестия слой. Невритите на тези клетки дават колатерали на гранулирания слой и отиват в
бели кахъри. Глиоцити. Кората на малкия мозък съдържа различни глиални елементи. Гранулираният слой съдържа фиброзни и протоплазмени астроцити. Дръжките на фиброзните астроцитни процеси образуват периваскуларни мембрани. Всички слоеве в малкия мозък съдържат олигодендроцити. Гранулираният слой и бялото вещество на малкия мозък са особено богати на тези клетки. Глиалните клетки с тъмни ядра лежат в ганглиозния слой между невроните с крушовидна форма. Процесите на тези клетки се изпращат на повърхността на кората и образуват глиални влакна на молекулярния слой на малкия мозък. Междуневронни връзки. Аферентните влакна, влизащи в кората на малкия мозък, са представени от два вида - мъхести и така наречените катерещи влакна. Мъхестите влакна преминават като част от маслиново-мозъчния и церебелопонтинния тракт и индиректно чрез гранулираните клетки имат възбуждащ ефект върху крушовидните клетки.
Катерещите влакна навлизат в кората на малкия мозък, очевидно, по дорзално-мозъчните и вестибулоцеребеларните пътища. Те пресичат гранулирания слой, прилепват към крушовидни неврони и се разпространяват по техните дендрити, завършвайки на повърхността им със синапси. Катерещите се влакна предават възбуждането директно към пириформените неврони.
Автономна (вегетативна) нервна система. Общи морфофункционални характеристики. отдели. Структура на екстрамуралните и интрамуралните ганглии.
ВНС е разделена на симпатикова и парасимпатикова. И двете системи участват едновременно в инервацията на органите и оказват противоположно въздействие върху тях. Състои се от централните части, представени от ядрата на сивото вещество на главния и гръбначния мозък, и периферните: нервни стволове, възли (ганглии) и плексуси.
Поради тяхната висока автономност, сложността на организацията и характеристиките на медиаторния метаболизъм, интрамуралните ганглии и пътищата, свързани с тях, се обособяват в независим метасимпатиков отдел на автономната НС. Има три типа неврони:
Еферентни неврони с дълъг аксон (клетки на Догел тип I) с къси дендрити и дълъг аксон, простиращ се отвъд възела до клетките на работния орган, върху който образува двигателни или секреторни окончания.
Аферентните неврони с еднакъв израстък (клетки на Догел тип II) съдържат дълги дендрити и аксон, който се простира отвъд този ганглий в съседните и образува синапси върху клетки от тип I и III. Те са част от локалните рефлексни дъги като рецепторна връзка, които са затворени без нервен импулс да постъпва в централната нервна система.
Асоциативните клетки (тип III Dogel клетки) са локални интеркаларни неврони, които свързват няколко клетки от тип I и II с техните процеси. Дендритите на тези клетки не излизат извън възела, а аксоните отиват към други възли, образувайки синапси върху клетки от тип I.
В автономната нервна системаправи разлика между централни и периферни региони. Централните отдели на симпатиковата нервна система са представени от ядрата на страничните рога на тораколумбалния отдел на гръбначния мозък. В парасимпатиковата нервна система централните отдели включват ядрата на средния и продълговатия мозък, както и ядрата на страничните рога на сакралния гръбначен мозък. Парасимпатиковите влакна на краниобулбарната област излизат като част от III, VII, IX и X двойки черепни нерви.
Периферни отдели на автономната нервна системаобразувани от нервни стволове, ганглии и плексуси.
Автономни рефлексни дъгизапочват с чувствителен неврон, чието тяло лежи в гръбначния възел (ганглий), както и в соматичната рефлексна дъга. Асоциативните неврони се намират в страничните рога на гръбначния мозък. Тук нервните импулси преминават към междинни преганглионарни неврони, чиито процеси напускат централните ядра и достигат до автономните ганглии, където предават импулси към моторния неврон. В тази връзка нервните влакна се разграничават преганглионарни и постганглионарни. Първият от тях напуска централната нервна система като част от вентралните корени на гръбначните нерви и черепните нерви. Както в симпатиковата, така и в парасимпатиковата система преганглионарните нервни влакна принадлежат към холинергичните неврони. Аксоните на невроните, разположени във автономните ганглии, се наричат постганглионарни. Те не образуват директен контакт с ефекторните клетки. Крайните им участъци по хода си образуват разширения - разширени вени, които включват медиаторни везикули. В областта на разширените вени няма глиална мембрана и невротрансмитерът, освободен в околната среда, засяга ефекторните клетки (например жлезисти клетки, гладки миоцити и др.).
в периферните ганглиисимпатиковата нервна система, като правило, са адренергични еферентни неврони (с изключение на неврони, които имат синаптични връзки с потните жлези, където симпатиковите неврони са холинергични). В парасимпатиковите ганглии еферентните неврони винаги са холинергични.
ганглииса клъстери от мултиполярни неврони (от няколко клетки до десетки хиляди). Екстраорганните (симпатикови) ганглии имат добре изразена съединителнотъканна капсула, като продължение на периневриума. Парасимпатиковите ганглии обикновено се намират в интрамуралните нервни плексуси. Ганглиите на интрамуралните плексуси, подобно на други автономни възли, съдържат автономни неврони на локални рефлексни дъги. Мултиполярните неврони с диаметър 20-35 μm са разположени дифузно, всеки неврон е заобиколен от ганглийни глиоцити. Освен това са описани невроендокринни, хеморецепторни, биполярни и, при някои гръбначни, униполярни неврони. В симпатиковите ганглии има малки интензивно флуоресцентни клетки (MYF клетки) с къси процеси и голям брой гранулирани везикули в цитоплазмата. Те секретират катехоламини и имат инхибиторен ефект върху предаването на импулси от преганглионарните нервни влакна към еферентния симпатиков неврон. Тези клетки се наричат интерневрони.
Сред големи мултиполярни неврониразграничават се вегетативни ганглии: моторни (тип I-тип Dogel клетки), чувствителни (тип II-тип Dogel клетки) и асоциативни (тип III-тип Dogel клетки). Моторните неврони имат къси дендрити с ламеларни разширения („рецептивни подложки“). Аксонът на тези клетки е много дълъг, надхвърля ганглия като част от постганглионарните тънки немиелинизирани нервни влакна и завършва на гладки миоцити на вътрешните органи. Клетките от тип I се наричат неврони с дълъг аксон. Неврони от II тип - равноотдалечени нервни клетки. От тялото им излизат 2-4 процеса, сред които е трудно да се различи аксон. Без да се разклоняват, процесите отиват далеч от тялото на неврона. Техните дендрити имат чувствителни нервни окончания, а аксонът завършва върху телата на двигателните неврони в съседните ганглии. Клетките тип II са чувствителни неврони на локалните автономни рефлексни дъги. Тип III Dogel клетки са подобни по форма на тялото на тип II автономни неврони, но техните дендрити не се простират отвъд ганглия и невритът отива към други ганглии. Много изследователи смятат тези клетки за разновидности на чувствителни неврони.
По този начин, в периферни автономни ганглииима локални рефлексни дъги, състоящи се от сензорни, двигателни и евентуално асоциативни автономни неврони.
Интрамурални автономни ганглиив стената на храносмилателния тракт се различават по това, че в техния състав, в допълнение към моторните холинергични неврони, има инхибиторни неврони. Те са представени от адренергични и пуринергични нервни клетки. В последния медиаторът е пуринов нуклеотид. В интрамуралните автономни ганглии има и пептидергични неврони, които секретират вазоинтестинален пептид, соматостатин и редица други пептиди, с помощта на които се извършва невроендокринна регулация и модулиране на активността на тъканите и органите на храносмилателната система.
Образователно видео за анатомията на автономната нервна система (ВНС)
В случай на проблеми с гледането, изтеглете видеоклипа от страницатаМускулните тъкани са тъкани, които се различават по структура и произход, но имат обща способност да се съкращават. Те се състоят от миоцити - клетки, които могат да възприемат нервните импулси и да реагират на тях със свиване.
Свойства и видове мускулна тъкан
Морфологични характеристики:
- Удължена форма на миоцити;
- надлъжно разположени миофибрили и миофиламенти;
- митохондриите са разположени в близост до контрактилни елементи;
- присъстват полизахариди, липиди и миоглобин.
Свойства на мускулната тъкан:
- контрактилност;
- възбудимост;
- проводимост;
- разтегливост;
- еластичност.
Има следните видовемускулна тъкан в зависимост от морфофункционалните характеристики:
- Набраздени: скелетни, сърдечни.
- Гладка.
Хистогенетична класификацияразделя мускулната тъкан на пет типа в зависимост от ембрионалния източник:
- Мезенхимен - дезмален зародиш;
- епидермална - кожна ектодерма;
- neural - неврална пластина;
- целомични - спланхнотоми;
- соматични - миотом.
От 1-3 вида се развиват гладкомускулни тъкани, 4, 5 дават набраздени мускули.
Структурата и функцията на гладката мускулна тъкан
Състои се от отделни малки вретеновидни клетки. Тези клетки имат едно ядро и тънки миофибрили, които се простират от единия край на клетката до другия. Гладките мускулни клетки са комбинирани в снопове, състоящи се от 10-12 клетки. Тази връзка възниква поради особеностите на инервацията на гладките мускули и улеснява преминаването на нервен импулс към цялата група гладкомускулни клетки. Гладката мускулна тъкан се свива ритмично, бавно и продължително, като същевременно е в състояние да развие голяма сила без значителен разход на енергия и без умора.
При нисшите многоклетъчни животни всички мускули са изградени от гладка мускулна тъкан, докато при гръбначните тя е част от вътрешните органи (с изключение на сърцето).
Контракциите на тези мускули не зависят от волята на човека, тоест възникват неволно.
Функции на гладката мускулна тъкан:
- Поддържане на стабилно налягане в кухи органи;
- регулиране на кръвното налягане;
- перисталтика на храносмилателния тракт, движение на съдържанието по него;
- изпразване на пикочния мехур.
Структурата и функцията на скелетната мускулна тъкан

Състои се от дълги и дебели влакна с дължина 10-12 см. Скелетните мускули се характеризират с доброволно свиване (в отговор на импулси, идващи от кората на главния мозък). Скоростта на съкращението му е 10-25 пъти по-висока, отколкото в гладкомускулната тъкан.
Мускулното влакно на набраздената тъкан е покрито с обвивка - сарколема. Под мембраната е цитоплазмата с голям брой ядра, разположени по периферията на цитоплазмата, и контрактилни нишки - миофибрили. Миофибрилата се състои от последователно редуващи се тъмни и светли области (дискове) с различен коефициент на пречупване на светлината. С помощта на електронен микроскоп беше установено, че миофибрилата се състои от протофибрили. Тънките протофибрили са изградени от белтък - актин, а по-дебелите - от миозин.
При свиването на влакната възниква възбуждане на контрактилни протеини, тънки протофибрили се плъзгат върху дебели. Актинът реагира с миозина, за да образува единна актомиозинова система.
Функции на скелетната мускулна тъкан:
- Динамично - движение в пространството;
- статично - поддържане на определено положение на частите на тялото;
- рецептор - проприорецептори, които възприемат дразнене;
- отлагане - течност, минерали, кислород, хранителни вещества;
- терморегулация - отпускане на мускулите с повишаване на температурата за разширяване на кръвоносните съдове;
- изражения на лицето - за предаване на емоции.
Структурата и функцията на сърдечната мускулна тъкан
 сърдечна мускулна тъкан
сърдечна мускулна тъкан Миокардът е изграден от сърдечен мускул и съединителна тъкан, със съдове и нерви. Мускулната тъкан се отнася до набраздените мускули, чиято набразденост също се дължи на наличието на различни видове миофиламенти. Миокардът е изграден от влакна, които са свързани помежду си и образуват мрежа. Тези влакна включват единични или двуядрени клетки, които са подредени във верига. Те се наричат контрактилни кардиомиоцити.
Контрактилните кардиомиоцити са с дължина от 50 до 120 микрона и ширина до 20 микрона. Ядрото тук се намира в центъра на цитоплазмата, за разлика от ядрата на набраздените влакна. Кардиомиоцитите имат повече саркоплазма и по-малко миофибрили от скелетните мускули. В клетките на сърдечния мускул има много митохондрии, тъй като непрекъснатите сърдечни удари изискват много енергия.
Вторият тип миокардни клетки са проводими кардиомиоцити, които образуват проводната система на сърцето. Проводимите миоцити осигуряват предаване на импулси към контрактилните мускулни клетки.
Функции на сърдечната мускулна тъкан:
- Помпена къща;
- осигурява притока на кръв в кръвния поток.
Компоненти на контрактилната система
Структурните характеристики на мускулната тъкан се определят от изпълняваните функции, способността за приемане и провеждане на импулси и способността за свиване. Механизмът на свиване се състои в координираната работа на редица елементи: миофибрили, контрактилни протеини, митохондрии, миоглобин.
В цитоплазмата на мускулните клетки има специални контрактилни нишки - миофибрили, свиването на които е възможно с приятелската работа на протеини - актин и миозин, както и с участието на Ca йони. Митохондриите захранват всички процеси с енергия. Също така енергийните резерви образуват гликоген и липиди. Миоглобинът е необходим за свързването на O 2 и образуването на неговия резерв за периода на мускулно съкращение, тъй като по време на свиване има компресия на кръвоносните съдове и доставката на O 2 към мускулите рязко намалява.
Таблица. Съответствие между характеристиките на мускулната тъкан и нейния тип
| Вид плат | Характеристика |
|---|---|
| гладък мускул | Включен в стените на кръвоносните съдове |
| Структурна единица - гладък миоцит | |
| Намалява бавно, несъзнателно | |
| Няма напречна ивица | |
| Скелетен | Структурна единица - многоядрени мускулни влакна |
| Характеризира се с напречна ивица | |
| Намалява бързо, съзнателно |
Къде се намира мускулната тъкан?
Гладките мускули са неразделна част от стените на вътрешните органи: стомашно-чревния тракт, пикочно-половата система и кръвоносните съдове. Те са част от капсулата на далака, кожата, сфинктера на зеницата.
Скелетните мускули заемат около 40% от телесното тегло на човек, с помощта на сухожилията те са прикрепени към костите. Тази тъкан се състои от скелетни мускули, мускули на устата, езика, фаринкса, ларинкса, горната част на хранопровода, диафрагмата, мимическите мускули. Също така, набраздения мускул се намира в миокарда.
Как се различават скелетните мускулни влакна от гладката мускулна тъкан?
Влакната на набраздените мускули са много по-дълги (до 12 cm) от клетъчните елементи на гладката мускулна тъкан (0,05-0,4 mm). Освен това скелетните влакна имат напречна набразденост поради специалното разположение на актиновите и миозиновите нишки. Това не е така за гладките мускули.
В мускулните влакна има много ядра, а свиването на влакната е силно, бързо и съзнателно. За разлика от гладките мускули, клетките на гладката мускулна тъкан са мононуклеарни, способни да се свиват бавно и несъзнателно.





